小型CRISPR/Cas9ベクターの開発とイネゲノム編集への利用
要約
Cas9、sgRNAをコードするDNA配列を直接連結し、一つのRNAポリメラーゼIIプロモーターで転写させた場合においても、機能的なCas9、sgRNAが生成され、その過程には内在RNaseおよびCas9が関与していることを明らかにする。
- キーワード:CRISPR/Cas9、ゲノム編集、リボザイム
- 担当:生物機能利用研究部門・遺伝子利用基盤研究領域・先進作物ゲノム改変ユニット
- 代表連絡先:電話029-838-8450
- 分類:研究成果情報
背景・ねらい
Cas9、sgRNA発現コンストラクトの小型化ならびに両者の発現部位を揃えることを目的に、Cas9とsgRNAをコードする配列を連結して一つのRNAポリメラーゼII(pol II)プロモーターにより転写させるゲノム編集方法が報告されている。この際、Cas9とsgRNA間にribozymeやtRNA、Csy4等の配列を配することでCas9、sgRNAをコードするRNAを積極的に切断しているが、切断位置によってはsgRNAに余計な配列が付加されることもある。本研究では、Cas9、sgRNA配列をそのまま連結した場合でも、機能的なCas9、sgRNAが生成されることを見出し、そのメカニズムについて解析を行う。
成果の内容・特徴
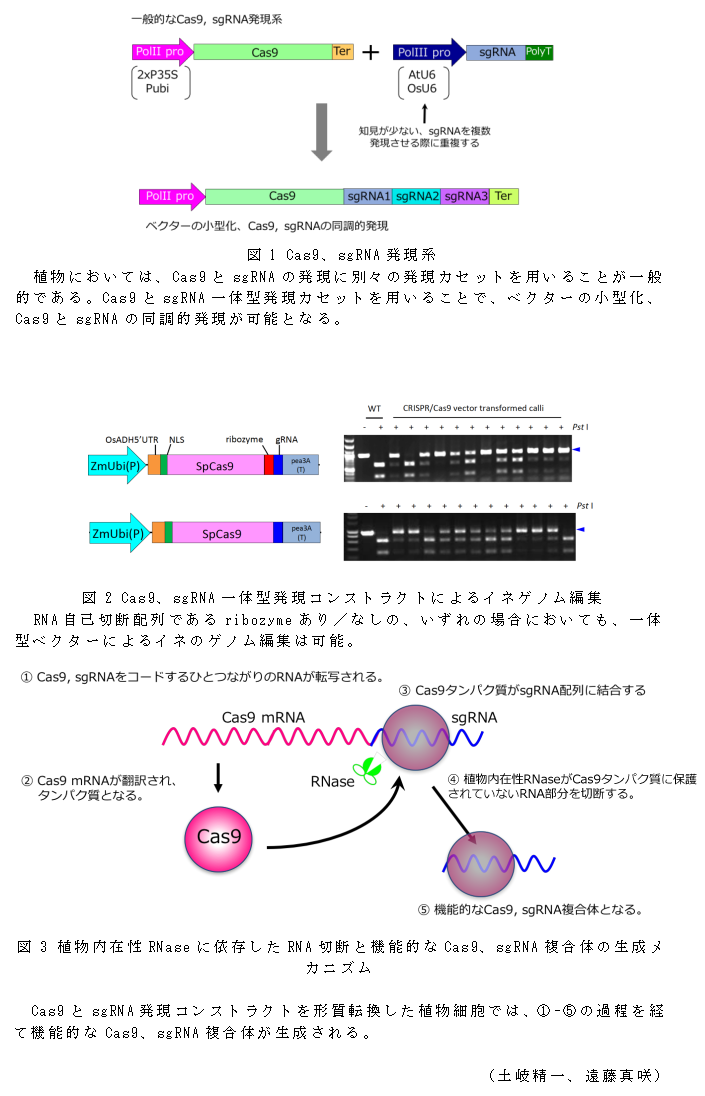
- 現在植物で主流となっているのは、Cas9、sgRNAを異なるプロモーター、ターミネーターで発現させる方法であるが、一つのプロモーターを用いて両者を発現させる一体型ベクターを用いることによりベクターの小型化、Cas9とsgRNAの同調的発現が可能になる(図1)
- Cas9、sgRNA一体型ベクターを用いてイネのゲノム編集を行う。Cas9、sgRNAをコードするRNAを分割する仕掛けとして、RNA自己切断配列であるリボザイム配列を含むベクター、含まないベクター、いずれにおいてもイネのゲノム編集は可能である(図2)。
- リボザイムを含まないベクターにおいても機能的なsgRNAが生成されたメカニズムについて以下のように考える。Cas9、sgRNAをコードするひとつながりのRNAが転写された後、Cas9は翻訳されてタンパク質となり、Cas9タンパク質がsgRNA配列に結合したのち、植物内在性RNaseがCas9タンパク質に保護されていないRNA部分を切断することで機能的なCas9、sgRNA複合体が生成される(図3)。
成果の活用面・留意点
- Pol II pro::Cas9::sgRNA(s)::Terは最小のCas9、sgRNAコンストラクトであり、形質転換効率を考えた場合は有利となる。
- 一方、イネにおける変異導入効率は、sgRNAを単独のU6プロモーターで発現させた場合の方が高いので、変異導入に適したCas9、sgRNAコンストラクトは植物種や、発現させたいsgRNAの数によって異なる。
具体的データ
その他
- 予算区分:交付金、その他外部資金(SIP)
- 研究期間:2015~2017年度
- 研究担当者:土岐精一、遠藤真咲、三上雅史(横浜市立大院)
- 発表論文等:Mikami M. et al. (2017) Plant Cell Physiol. 58(11):1857-1867