イネの花器官形成を制御する新規MADSボックス転写因子
|
|
| [要約] |
イネの mfo1 変異体は花器官形成にさまざまな異常を生じる。原因遺伝子である MFO1 は、これまで機能不明であったAGL6-like MADSボックス転写因子をコードし、内穎周縁部、鱗被、胚珠の形成を制御するとともに穎花形成の繰り返しを抑制している。 |
![]()
[キーワード]イネ、突然変異、花器官、形態形成、転写因子、MADSボックス、 MFO1 |
![]()
[担当]中央農研・稲遺伝子技術研究北陸サブチーム
[代表連絡先]電話:025-526-8251
[区分]作物、関東東海北陸農業・北陸・水田作畑作
[分類]研究・普及 |
|
| [背景・ねらい] |
遺伝子拡散抑制技術として期待される閉花受粉性イネ「spw1-cls」の原因遺伝子 SPW1 は花器官形成を制御するMADSボックス遺伝子群のひとつで、鱗被(穎を押し広げて開花を引き起こす花器官)およびおしべの形態形成を制御している。閉花受粉性を安定的に利用するためには、SPW1と協調して花器官形成を制御している遺伝子群の機能を理解することが重要である。そこで本研究では、鱗被などの花器官形成に異常を示すイネの突然変異体の解析によって、花器官形成を制御する新規遺伝子の機能を明らかにする。 |
![]()
[成果の内容・特徴] |
| 1. |
イネ変異体mosaic floral organs1 (mfo1)は、野生型(図1A,B)と比較して、内穎周縁部の消失(内穎の外穎化)、鱗被の穎化、おしべの減少、胚珠の発達異常などの形態異常を示す(図1C,D)。さらに mfo1 では、穎花が繰り返し形成される(図1E)。 |
| 2. |
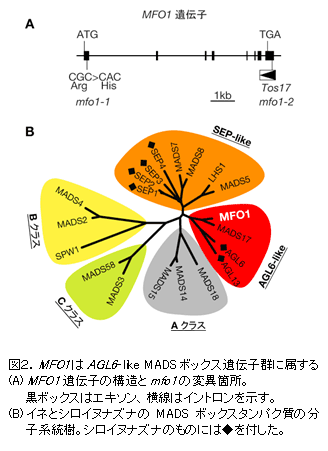
mfo1 には2つの変異型アリル(対立遺伝子型)がある。強い形態異常を示す mfo1-1 アリルでは、AGL6-like MADSボックス遺伝子内の塩基置換でアミノ酸置換を生じている。弱い形態異常を示す mfo1-2 アリルでは、3’非翻訳領域内へのレトロトランスポゾン Tos17 の挿入により、同遺伝子の発現量が低下している。したがって、これら mfo1 変異体の原因遺伝子 MFO1 はAGL6-like MADSボックス遺伝子と結論できる(図2A)。 |
| 3. |
MADSボックス遺伝子群は、塩基配列から推定されるアミノ酸配列により、A、B、C、D、SEP-like、AGL6-likeなどのクラスに分類できる。 MFO1 遺伝子は、イネのMADS17やシロイヌナズナのAGL6、AGL13等の遺伝子とともに、これまで機能未知であったAGL6-like遺伝子群に分類される(図2B)。 |
| 4. |
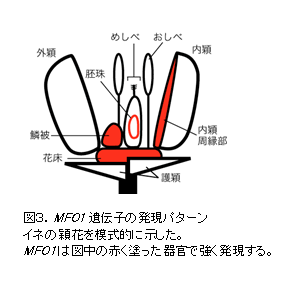
MFO1 は花床、内穎周縁部、鱗被、胚珠の珠皮などで強く発現する(図3)。これらの発現領域は、 mfo1 変異体で形態異常が生じる領域と一致する。 |
| 5. |
LHS1 遺伝子は SEP-likeに分類され、花器官形成に異常を示すlhs1-2 変異体(図1F)の原因遺伝子として知られている。 mfo1-2 と lhs1-2 の二重変異体では、内部花器官が形成されず、しばしば内部に新たな穎花が繰り返して形成される(図1G,H)。 |
| 6. |
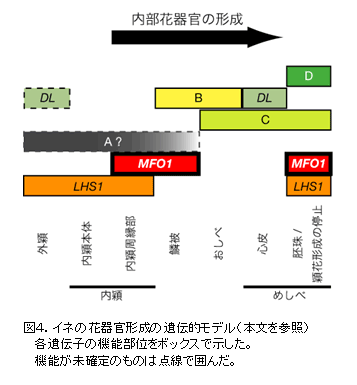
これまで提唱されていたイネの花器官形成モデル(「ABCモデル」)中に、今回解明した MFO1 の役割を当てはめた改訂モデルを図4に示す。 MFO1 は、SPW1を含むBクラスとともに鱗被形成を、また LHS1 とともに内穎周縁部形成を制御する。また MFO1 は、Cクラス、Dクラスおよび LHS1 とともに胚珠形成を制御する。さらに MFO1 は LHS1 とともに、内部花器官形成を誘導し、穎花形成の繰り返しを抑制する。 |
|
![]()
[成果の活用面・留意点] |
| 1. |
植物の花器官形成の分子機構を解明するための重要な知見である。 |
| 2. |
閉花受粉性イネの安定的利用技術の開発をはじめ、作物の花器官の形態を改変するための知見として利用できる。 |
|
![]()
[具体的データ] |
|
| [その他] |
研究課題名:遺伝子組換え技術の高度化と複合病害抵抗性等有用組換え稲の開発
中課題整理番号: 221h
予算区分:委託プロ(新農業展開)、交付金プロ(形態・生理、組換え植物)
研究期間:2004〜2009年度
| 研究担当者: |
大森伸之介、吉田均、木水真由美、杉田麻衣子(東大)、宮尾安藝雄(生物研)、内田英史、廣近洋彦(生物研)、長戸康郎(東大) |
発表論文等:Ohmori et al. (2009) Plant Cell 21(10): 3008-3025
|
|
| 目次へ戻る |