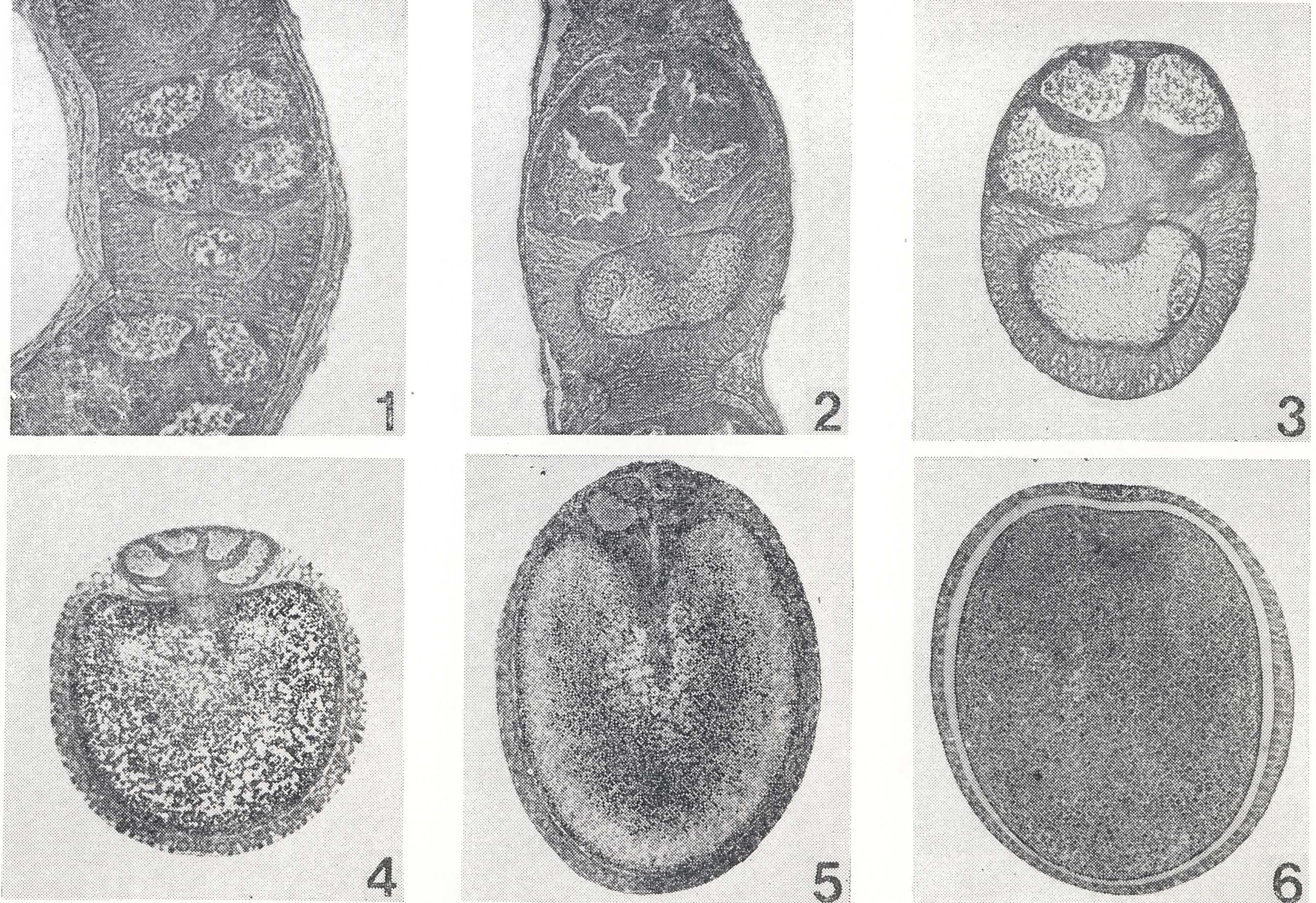
第16図 卵母細胞の発育階程。数字は階程番号(本文参照)
Ⅲ 上蔟・種繭保護と造卵および産卵
蚕は、普通、上蔟後3日ぐらいで吐糸を終了し、脱皮して蛹になるから、上蔟後7、8日で収繭し、蚕種製造所に持込むまでには蛹期間の約3分1が経過している。蚕が上蔟して繭を作ってしまうと、農家の蚕に対する注意は、飼育中に比べておろそかになりがちである。一般に、春蚕では上蔟後の補温に欠けることが多いが、また反対に、補温したまま温度が上り過ぎても気付かないことがある。
吐糸終了から蛹の初期にかけては、生理的にも形態的にも蚕の体に重大な転換の行なわれる時期であるから、この時期の管理がなおざりにされていることは蚕種製造技術上の一つの盲点であって、不受精卵が多発した場合にその原因を調べようとしても、製造所に入ってからの保護環境はわかるが、それ以前のことは不明な場合が多い。
糸繭の目的である繭層は上蔟後3目ぐらいででき上るが、種繭の目的である卵の造成の重要な部分は吐糸終了後に始まるのであるから、農家ばかりでなく、蚕種製造者自身が十分な注意を払う必要がある。普通に種繭保護と云えば収繭後の保護であるが、実際問題としては、これだけでは不十分である。
1 上蔟・吐糸
繭層は種繭にとって最終の目的物ではないが、吐糸が十分に行なわれず体内に絹物質が残留すると、造卵数も産卵数も少なくなるから(Ⅱ1B)、吐糸・営繭の環境に注意する必要がある。多糸量系の原種には一般に残留絹が多いと云われている。過熟、若上げは避けなければならない。
繭の縦作りがよいか横作りがよいかについては、古くから論議があり、昆虫には本来縦作りの性質があるから、この本来の性質に合わせるべきであると云う説もあるが、種繭としては縦作りよりも横作りが望ましいとされている。これは、縦作りにおいては、蚕が吐糸を終えて化蛹する際に、頭部を上に向けるので、尾部が圧縮されて交尾不能の蛾や離交し易い蛾ができるばかりでなく、上蔟後の斃蚕歩合、発蛾歩合、造卵数および産卵数などにも悪影響があると云われているからである(第34表)。
| 営繭姿態 | 発蛾歩合 (%) |
蛹体尾部異常 蚕歩合 (%) |
正常蚕卵蛾 歩合(%) |
採種総蛾数による1蛾当り | 種繭1,000粒当り正常卵 | |||
| 産卵数(粒) | 不良卵歩合(%) | 造卵数(粒) | 重量(g) | 指数 | ||||
| 横 斜 縦 |
86 76 80 |
9 74 94 |
93 74 67 |
455 415 390 |
2 4 5 |
485 473 475 |
108.8 85.8 78.0 |
100 79 72 |
池田(241)は、種々な向きに繭を作らせて実験的にこれを確かめ、全部の蚕が横作りの繭を作るような廻転蔟の使用をすすめている。ただ、池田の実験は4カ年、6蚕期に亘って繰返えされてはいるが、全体を通じて未化蛹斃蚕歩合や斃蛹歩合が非常に高く、第34表の場合(昭和19年秋)には、未化蛹斃蚕歩合が横作りにおいて35.6%、縦作りにおいて48.4%にも達している。蚕の不健康なとき、縦作りと横作りとで斃蚕歩合の差の大きいときなどに卵に関する成績の差も大きく出ているようにみえるから、蚕の健康なときには産卵に対してこれほど大きな差はないものと思われる。
加藤ら(314)は、豊白、研光、支124号、銀竜および高の5品種についての比較を行なったが、造卵数は、支124号において縦作りが37.9±8.1粒少なかっただけで、他の品種においては縦作りと横作りとの間に差がなかった。発蛾歩合、正常産卵蛾歩合にも一定の傾向の違いは認められなかった(Ⅲ5)。
吐糸中の蚕は繭中で体の位置を種々に変えているが、吐糸を終えると繭の一端に頭部を向けて静止し、化蛹する。この静止するときの頭の向きは重力と光とによって強く支配され、水平に営繭させて一方向から光をあてると、光の来る方向に頭部を向けているものが、反対に向いているものより多い。しかし、光よりも重力の影響が強いので、傾斜した位置で営繭させると、光の方向とは無関係に大部分または全部のものが上方に頭部を向ける(第35表)。
| 営繭角度 | 光線 | 繭内の蛹の頭部の向き | |||
| N品種 | S品種 | ||||
| 上(頭) | 下(頭) | 上(頭) | 下(頭) | ||
| 15° | 全暗 上明、下暗 上暗、下明 |
28 28 27 |
2 2 3 |
23 27 19 |
7 3 11 |
| 30° | 全暗 上明、下暗 上暗、下明 |
30 30 30 |
0 0 0 |
29 30 25 |
1 0 5 |
| 45° | 全暗 上暗、下明 |
30 30 |
0 0 |
30 29 |
0 1 |
| 90° | 全暗 上暗、下明 |
30 30 |
0 0 |
30 29 |
0 1 |
但し、品種によって反応にかなり相違がある。また、水平に営繭させた繭の蛹の尾端に当る方の破風部に250luxの光をあてると、吐糸終了後5時間(上蔟後約63時間)では約25%の蚕が光の方向へ頭部の向きを変えたが、吐糸終了後10時間(上蔟後約68時間)では光に反応して向きを変えるものはなかった。これに対し、暗中において45°の角度で営繭させたものの上下を反対にすると(45°の傾斜で)、吐糸終了後5時間では約半分、10時間でもなお約22%の蚕が上方へ向きを変えた。即ち、250luxの光よりも45°の角度による重力の刺戟の方が強く(おそくまで)働いている(422)。これは、供試品種33×73、温度25℃ 、湿度約80%、上蔟後約20時間で営繭中の幼虫が僅かに透視できる程度に吐糸し、その後38時間ぐらいで吐糸終了、その後更に35時間で化蛹した場合における調査結果である。
池田(241)は、上蔟後3日間を第1期、次ぎの3日間を第2期、その後発蛾までを第3期として、繭の向きを変える実験を行ない、頭部を上方に向けて第2期(吐糸終了から若蛹の期間)を径過させると前記の縦作りの被害が最も著しいと云っている。縦作りによる尾部の圧縮障害は、第2期の初めに(欧16号では上蔟後102時間目)に上下を逆転すれば防止できる(小泉の結果からも理解できる)が、斃蚕を多くしたり発蛾歩合、正常産卵蛾歩合および造卵数などを低下させる生理障害は、この時期に繭を逆転しても防止できなかった。
吐糸能率を高め、残絹量を少なくするための蔟中温度は、川畑(323)によれば23℃で、25.5℃、23℃゜、20℃における残絹量はそれぞれ13%、4%および6%であった。乾繰上蔟(湿度75%)に比べて多湿上蔟(90%)、有気流上蔟(風速1.0m/sec)に比べて無気流上蔟はそれぞれ残留絹が幾分多いと云うから、換気のよい乾燥状態で上蔟させることも必要である。
2 卵母細胞および蛹の発育階程
蚕種の保護が胚の発育に基礎をおいているのと同様に、種繭の保護も卵母細胞および蛹の発育に基ずき、その生理に応じた取扱いをする必要がある。
卵母細胞の発育には、胚の場合のような一般的に採用されている基準階程がないが、小沢(720)は、実用のために七つの発育階程を区別した(第16図)。
第16図 卵母細胞の発育階程。数字は階程番号(本文参照)
1 初期球状期 栄養細胞と卵母細胞との配列が交互栄養型(Ⅰ2Aa)の特徴を現わした時期で、卵母細胞は球状であるが、栄養室側が平たく、下方に向ってまるい。その核は細胞の中央に位し、球形である。核内に大形のまるい仁がみられる。包卵細胞は未だ一定の配列を示さず、卵母細胞の外側に散在する。
2 盃状期 栄養細胞の発育が進み、卵母細胞は、これに接する側が圧迫されて、盃状を呈する。卵母細胞の核は片側に寄り、仁は染色性が淡くなる。包卵細胞は卵室外側にほぼ1列にならび、次第にこれを取囲む。栄養細胞とその上位の卵母細胞との間に橋帯ができる。
3 半球状期 包卵被膜がほぼ完成し、これに包まれた卵母細胞が濾胞のなかばを占めて半球状を呈する時期である。卵母細胞に淡黄色の脂肪性卵黄が蓄積されるために、生の材料で核を透視することができなくなる。卵母細胞の中央部に、栄養細胞からの大量の物質の流入が認められる。卵母細胞を取囲む包卵被膜細胞は柱状の丈の高い細胞になる。栄養細胞と卵母細胞との間にも、周囲から包卵細胞が入込んでくるが、なお巾の広い連絡が残っている。
4 栄養細胞退化期 卵母細胞は縦の方向に長さを増し、上部の大きく欠けた楕円形になる。栄養細胞と卵母細胞とはなお連絡し、前者から後者への物質の流入が認められる。
5 包卵被膜完成期卵 栄養細胞と卵母細胞との間に入込んだ包卵被膜細胞が発達して、卵室を完全に取囲む時期である。卵母細胞は完全な楕円形に近付き、上部に僅かに凹みを残すが、大きさはほぼ成熟卵に等しい。卵黄膜と包卵被膜との開に卵殻が形成され初める。
6 卵殻分泌期 包卵被膜細胞から卵殻の分泌される時期である。
7 完成期 卵管本部の卵が包卵被膜から離脱して下降する時期である。
蛹令別に、1卵管中に含まれる卵をこれらの階程によって示したのが第36表であるが、これに勝又(317)の蛹の発育階程を併記した。一般に、複眼が黒色になったときは、その保護条件における蛹期間のなかばを幾分過ぎた時期とされている。
| 蛹令 | 調査 卵管数 |
卵母細胞の発育階程 | 蛹の外形 | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 産下卵 | |||
| 化蛹 1日目 |
本 |
粒 |
粒 |
粒 |
粒 |
粒 |
粒 |
粒 |
粒 |
体柔らかく黄白色または淡黄褐色。翅下の腹部 白色。頭部透明、正中腺および触角基部黒色に 透視される。 |
| 2日目 | 11 | 81 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 翅下の腹部褐色となる。 |
| 3日目 | 11 | 39 | 59 | 8 | 0 | 0 | 0 | 0 | 0 | 体硬化、固有色 |
| 4日目 | 12 | + | 30 | 22 | 26 | 0 | 0 | 0 | 0 | 尾部透明となる。 |
| 5日目 | 10 | + | 24 | 13 | 31 | 38 | 0 | 0 | 0 | 眼着色し始め、小豆色。 |
| 6日目 | 11 | + | 16 | 24 | 51 | 0 | 0 | 0 | 眼濃小豆色。 | |
| 7日目 | 10 | + | 13 | 38 | 28 | 0 | 0 | 眼黒色。頭部白色不透明。 | ||
| 8日目 | 11 | + | 18 | 21 | 53 | 0 | 0 | 生殖キチン板、爪、肢の環節 など黒色に透視される。 |
||
| 9日目 | 10 | + | 13 | 13 | 69 | 8 | 0 | 体銹色を帯び、触角着色し始む。 | ||
| 10日目 | 12 | + | 13 | 48 | 31 | 0 | 触角暗褐色、体銹色を増し、柔かし | |||
| 11日目 | 11 | + | 12 | 17 | 67 | 0 | 体柔かくなり、土色、皺多く、翅の斑紋みえるもの あり(みえないで発蛾することもある)。 |
|||
| 発 蛾 | 20 | + | 11 | 6 | 79 | 0 | ||||
| 産卵後 | 10 | + | + | 2 | 0 | 81 | ||||
このような蛹の形態変化は複雑な変態および発育の生理に件なって進行している訳であるが、生理作用の外面的な現われの一つである呼吸は蛹期間を通じて第17図のような消長を示す。蚕の酸素消費量は5令の中期以後低下を続け、化蛹3日目頃に最低になるのであるが(266)、伊藤(280)は、蛹体休眠をする鱗翅目昆虫の生理状態と比較して、この呼吸最低期は組織崩壊と組織形成との二つの異なる代謝の転換期に当り、生理的には休眠状態に近いものと考えている。
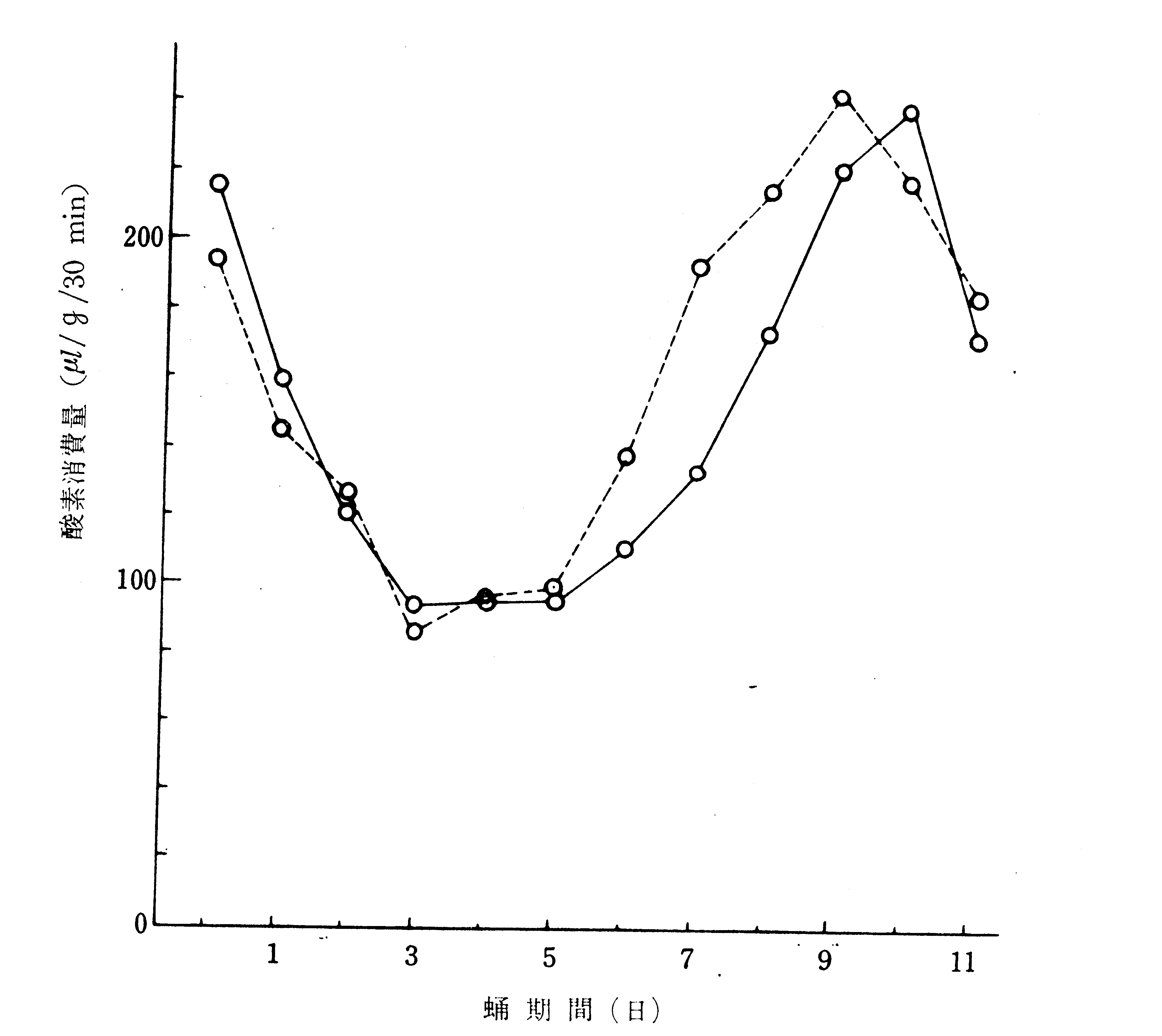
第17図 蛹期間における酸素消費量の消長(伊藤(280))
点線は雌、実線は雄
この考えの当否は兎も角、化蛹3日目頃の呼吸最低期を中心とする時期は、蛹の保謹上からも極めて重要な転換期である。この最低期を境に雌と雄との曲線の上下が入れかわることについて、高岡(958)は、卵巣を除去すると雌の曲線が雄型になることから、雌における卵巣の発育に伴なう酸素消費の増加が原因であろうと考えている。但し、高岡の酸素消費量曲線は、化蛹後約50時間目を最低として、雄はやや直線的に尻上りの増加を示し、雌は170時間目頃に最高となり以後減少すると云うから、第17図とは少しく異なるもののようである。後に述べるように、休眠ホルモンが卵に作用し初めるのはこの最低期である。蛹期中で冷蔵に最も耐えるのはこの時期であり、これに次ぐのが、第2の呼吸低下期に当たる化蛹前日であることも注目される。
木村・水出(379)によれば、化蛹後、温度25.3℃(23.5℃-27.5℃)、湿度53.6%(41-65%)で保護した場合(蛹期間は雌平均253時間、雄平均246時間)の蛹重減耗曲線には、化蛹後80時間目前後の、複眼が淡茶色に着色し初めたときに変曲点があり、
┌ W1=4.655 e-0.0258t1 +95.25 0≦t1≦80
雌 ┤
└ W2=-0.011210(t2-70)1.3564+96.05 80≦t2≦240
┌ W1=5.619 e-0.0280t1 +96.05 0≦t1≦80
雌 ┤
└ W2=-0.004914(t2-70)1.5864+94.83 80≦t2≦240
で表わせると云う。但し、t1、t2は化蛹後の経過時間、W1、W2は、t1、t2における蛹重(化蛹直後を100とする指数)である。品種はC110×N112であった。
3 蛹の保護
A 蛹の発育
a 保護環境と蛹期間
普通の保護温度の範囲内では多くの場合、雌よりも雄の蛹期間が短い。蛹期間は温度の上昇に連れて短かくなるが(第37表)、堀内ら(223)によれば、温度に正しく比例する訳ではなく、20℃と25℃との間では40%以上も短縮するが、25℃と30℃との間の短縮は10%余りに過ぎない。温度による蛹期間の変化は、また雌雄によって異なり、雌雄差は25℃において最大で、30℃、20℃および15℃においてはこれよりも差が小さい。特に、15℃においては、従来から知られているように雄よりも雌の蛹期間が短かくなる傾向がみられたが、30℃においても、幾分これと同じような傾向のあることがわかった(第40表)。 この関係は品種によっても異なる。
| 保護温度 ℃ |
性別 | 蛹 期 間 (日) | |||
| 日122号 | 日124号 | 支115号 | 支124号 | ||
| 30 | ♀ ♂ |
**10.6 10.0 |
** 9.6 9.3 |
* 8.0 7.7 |
** 8.3 7.8 |
| 25 | ♀ ♂ |
**11.9 11.1 |
**10.6 10.3 |
9.4 9.1 |
* 9.8 9.5 |
| 20 | ♀ ♂ |
**20.3 19.8 |
18.5 18.6 |
15.9 **16.9 |
16.7 16.7 |
| 15 | ♀ ♂ |
26.3 26.0 |
24.6 24.9 |
22.1 22.3 |
**23.0 22.4 |
上蔟は殆ど総べての品種において雄が雌より早いが、化蛾は必ずしもそうではなく、諸星・小林(599)によれば、個体別に化蛹時刻を調べ、24℃と19℃とで保護したものについて蛹期間の平均を求めると、両温度区を通じて、カムボージュ、欧16号、欧19号などにおいては雄が雌より短かく、大造、日112号などでは雌が雄より短かかった。温度との関係については、保護温度の高いほど雄の相対的発育速度が雌より大きくなり、温度が低いほど雌の相対的発育速度の大きくなる傾向がある。これは伴性晩性遺伝子の働きとホルモン作用の拮抗とによって説明できると云う(597)。永友(645)、室賀(617)も蛹期間の雌雄差についての遺伝学的説明を試みている。
堀内ら(223)の上記の実験は、6時から21時までを明保護、21時から翌朝6時までを暗保護にして行なったのである.が、6時に明るくすると大部分の蛾は3時間以内に発蛾した。その後にも不時発蛾するものがあるが、不時発蛾は、試験した温度の範囲内(第37表)では、25℃保護において最も少なかった。
保護温度の高低によって蛹期間が伸縮すると、蛹の外形の発育階程もこれに連れてほぼ均等に伸縮するもののようで、堀内らの試験の範囲内では、複眼小豆色のときが総ての保護温度を通じて蛹期間の中点であった。
蛹体の外観によって発蛾までの日数を推定することは古くから行なわれており、一般に、複眼が黒色になったときは蛹期のなかばを過ぎた時期であると云われているが、最近の品種についての加藤(311)の調査を第38表に示す。
| 蚕品種 | 日122号 | 秋花 | 支115号 | 支124号 | ||||||||
| 蛹の外観 | 22℃ | 25℃ | 28℃ | 22℃ | 25℃ | 28℃ | 22℃ | 25℃ | 28℃ | 22℃ | 25℃ | 28℃ |
| 上蔟 吐糸終了 化蛹 複眼着色始む 複眼淡黒色 複眼濃黒色 生殖外器着色始む 生殖外器黒色 生殖キチン板透視 触角濃黒色 蛹皮土色 |
19日 15 13 10 9 7 6 5 4 2 |
18日 15 13 10 9 7 6 5 4 2 |
15日 12 11 8 6 5 4 3 2 |
20日 15 14 11 7 6 5 3 2 |
17日 13 12 10 6 5 4 3 2 1 |
15日 12 11 9 5 4 3 2 1 |
16日 13 12 10 7 5 3 2 1 |
14日 11 10 8 6 4 3 2 1 |
13日 11 10 7 5 3 2 1 |
16日 13 12 9 6 4 3 2 1 |
16日 13 12 9 6 4 3 2 1 |
14日 11 10 8 6 4 3 2 1 |
保護湿度60-90%の範囲内では、湿度の低いほど蛹期間は短かく(第39表)、裸蛹は繭中のものより蛹期が短かいと云う。
| 湿 度 | 平均蟄繭日数* |
| 60% 70 80 90 |
13.2日 13.7 13.7 14.1 |
b 化蛹までの蔟中保護温度と蛹期間
化蛹後の保護温度と蛹期間との間に上記のような関係があるのに対し、化蛹までの蔟中保護温度を25℃にしたものと30℃にしたものとの蛹期間を比較すると、後者が前者よりも常に長いと云う(第40表)。
牛込(1127)は、日1号、日110号、支4号、支102号および欧7号について、上蔟以後の保護温度78-83゚F(25.6℃-28.3℃)の範囲内においては、温度の高いほど蛹期間が短縮するが、85゚F(29.2℃)また87゚F(30.6℃)がその限界で、保護濃度がそれ以上になると蛹期間は却って延長すると述べている。しかし、これは化蛹前および化蛹後を含めた期間であって、両者を区別した期間には言及していない。
| 化蛹後の保護温度 (℃) |
化蛹までの蔟中温度 (℃) |
性別 | 蛹期間(日) | |
| 日124号 | 支124号 | |||
| 30 | 25 | ♀ ♂ |
10.9 10.7 |
9.0 8.8 |
| 30 | ♀ ♂ |
11.0 *11.4 |
**9.4 8.7 |
|
| 25 | 25 | ♀ ♂ |
**13.7 12.6 |
**11.2 10.6 |
| 30 | ♀ ♂ |
13.9 13.4 |
**11.5 10.5 |
|
| 20 | 25 | ♀ ♂ |
20.8 20.9 |
**18.0 17.2 |
| 30 | ♀ ♂ |
21.3 21.2 |
**18.8 17.3 |
|
| 15 | 25 | ♀ ♂ |
30.5 30.5 |
**26.8 26.1 |
| 30 | ♀ ♂ |
31.9 31.0 |
**27.8 26.6 |
|
c 発育段階別保護温度と蛹の発育
伊与田・米山(295)は、蛹期の卵巣重にみられる3段階の発育(Ⅰ2Abi)は蛹の外形変化と関連があり、発育の前期は化蛹後、複眼の着色し初める時期まで、中期はその後触角の着色し初める時期までであるとし、低温(15℃)に保護すると前期と後期の延長は著しいが、中期の延長は比較的少ないと述べている(第41表)。
| 蛹令 (日目) |
第1区 全期高温 | 第2区 前期高温、中期 低温、後期高温 |
第3区 前期低温、 中・後期高温 |
第4区全期低温 | ||||||
| 蛹重 (g) |
卵巣重 歩合(%) |
蛹重 (g) |
卵巣重 歩合(%) |
保護 温度(℃) |
蛹重 (g) |
卵巣重 歩合(%) |
保護 温度(℃) |
蛹重 (g) |
卵巣重 歩合(%) |
|
| 化蛹当日 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 |
8.6 - 8.12) 8.23) 8.06) 7.78) 8.09) 8.110) 5.511) |
0.7 - 4.5 8.5 20.0 33.0 38.1 37.3 47.5 |
8.6 - 8.1 8.2 8.05) -6) - 8.17) - - - 8.1(5)9) - 4.711) |
0.7 - 4.5 8.5 13.2 - - 26.4 - - - 33.1 - 49.6 |
30 〃 〃 〃 15 〃 〃 〃 17 19 21 30 〃 〃 |
8.6 - - 8.6 - - 8.5 - - 8.3 7.96) 7.4(5) 7.6 4.911) |
0.7 - - 2.0 - - 3.3 - - 6.5 21.0 37.0 33.6 49.5 |
30 〃 〃 〃 15 〃 〃 〃 17 19 30 〃 〃 〃 |
8.6 - - 8.6 -1) - 8.5 - - 8.33) -4) - - 7.5 - - 7.99) - - 8.3(5) - 5.8(10)11) |
0.7 - - 2.0 - - 3.3 - - 6.5 - - - 31.7 - - 33.7 - - 31.2 - 46.1 |
加藤(311)は、全期間22℃、25℃および28℃の恒温区と、複眼黒色期(第38表の淡黒色期と濃黒色期とを合わせた時期)以後を変温した区とを設けて、上蔟後、発蛾までの保護を行ない、発蛾および産卵におよぼす温度の影響を比較した(第42表)。これによると、発蛾歩合は、上蔟後、発蛾までの全期間を同一温度で通した場合には28℃区が最も悪く、牛込(1127)の成績において、上蔟後を68゚F(20℃)、73゚F(22.8℃)、78゚F(25.6℃)および83゚F(28.3℃)に保護した場合に83゚F区の最も劣っていたことと一致した。22℃区と25℃区との成績は、品種的な関係があるためか、優劣をきめかねる結果であるが、複眼黒色期以前を22℃、以後を25℃に保護した区(表の22℃→25℃)が全期間25℃の区に勝っているのをみると、少なくとも発蛾歩合に関しては、化蛹前あるいは蛹初期の保護の適温は25℃以下のように考えられる。
| 蚕品種 | 保護温度 (℃) |
発蛾 歩合(%) |
正常産卵蛾 歩合(%) |
正常産卵蛾についての調査 | |||||
| 1蛾当り総 産卵数(粒) |
正常卵 歩合(%) |
卵長 (mm) |
卵巾 (mm) |
1g卵 数(粒) |
100粒 卵重(g) |
||||
| 日122号 | 22 25 28 22→25 22→28 25→22 28→22 |
88 83 67 86 78 83 70 |
86 86 57 76 76 88 47 |
558 542 462 612 525 517 526 |
94 96 87 96 96 95 92 |
1.29 1.29 1.33 1.30 1.29 1.31 1.34 |
1.04 1.05 1.07 1.05 1.04 1.06 1.09 |
1,731 1,728 1,591 1,698 1,683 1,685 1,637 |
0.0578 0.0579 0.0629 0.0589 0.0594 0.0594 0.0611 |
| 秋 花 | 22 25 28 22→25 22→28 25→22 28→22 |
77 63 55 70 68 68 66 |
70 69 47 55 40 51 40 |
511 494 457 497 514 468 445 |
97 95 78 96 96 92 73 |
1.36 1.34 1.37 1.33 1.33 1.34 1.36 |
1.10 1.09 1.10 1.08 1.08 1.08 1.11 |
1,676 1,667 1,615 1,671 1,646 1,680 1,617 |
0.0597 0.0600 0.0619 0.0598 0.0608 0.0595 0.0618 |
| 支115号 | 22 25 28 22→25 22→28 25→22 28→22 |
84 92 61 94 69 96 69 |
58 88 53 81 60 72 63 |
503 559 521 550 558 505 530 |
94 94 93 92 93 93 93 |
1.31 1.33 1.38 1.35 1.37 1.37 1.40 |
1.05 1.05 1.09 1.07 1.08 1.07 1.10 |
1,821 1,832 1,680 1,794 1,761 1,724 1,673 |
0.0549 0.0546 0.0595 0.0557 0.0568 0.0580 0.0598 |
| 支124号 | 22 25 28 22→25 22→28 25→22 28→22 |
90 91 74 92 73 92 62 |
66 88 53 68 65 73 60 |
520 563 536 612 503 565 469 |
93 94 88 92 92 94 90 |
1.27 1.28 1.30 1.27 1.26 1.29 1.29 |
1.07 1.07 1.08 1.06 1.05 1.08 1.09 |
1,770 1,768 1,700 1,787 1,706 1,714 1,657 |
0.0565 0.0566 0.0588 0.0560 0.0586 0.0583 0.0604 |
B 保護温度と造卵および産卵
a 造卵数・産卵数
普通の種繭保護温度の範囲内では、一般に、温度の低い場合に造卵数が多いが、第43表において、日110号および昭和A(生種)は20℃保護の造卵数が25℃あるいは30℃保護に比べて多いのに対し、昭和A(黒種)は20℃保護が25℃および30℃保護よりも却って造卵数が少ないのをみると、造卵数を増加させる低温の限度は品種または条件によって相違するものと考えられる。
| 保護温度 (℃) |
日110号 | 昭和A(黒種) | 昭和A(生種) | |||
| 調査蛾数 (蛾) |
1蛾平均 造卵数(粒) |
調査蛾数 (蛾) |
1蛾平均 造卵数(粒) |
調査蛾数 (蛾) |
1蛾平均 造卵数(粒) |
|
| 30 25 20 15 |
102 270 |
519 572 |
85 40 60 33 |
615 643 602 452 |
367 505 425 |
431 497 499 |
蛹の保護温度が造卵数の増加に影響する時期は、伊与田らによって、主として先に述べた前期と考えられている(第44表)。
| 蛹の保護条件 | 調査蛾数(蛾) | 平均造卵数(粒) |
| 全期間高温(第41表第1区) 前期高温、中期低温、後期高温(同表第2区) 前期低温、中・後期高温(同表第3区) 全期間低温(同表第4区) |
7 7 7 10 |
538 528 553 599 |
しかし、この造卵数の増加は卵巣の発育とは別で、卵巣の重量増加に最も大きく影響するのは中期の高温であると云う。
永井(630)が、支17号を65゚F(18.3℃)、75゚F(23.9℃)および90゚F(32.2℃)に分けて上蔟させ、化蛹後は何れも70゚F-72゚F(21.3℃-22.2℃)で保護した結果によると、卵管1本当りの完成卵数が65゚F区83.6粒、75゚F区94.4粒、90゚F区79.7粒で、75゚F区が最も多かった。これからみると、化蛹前の保護温度も造卵数に影響している。
造卵数は、このように、ある程度までは、蛹期の保護温度の低いほど増加するが、造成された卵の産卵歩合、正常産卵蛾歩合、不受精卵歩合などからみると、後に述べるように、低温保護の成績が不良で、卵の孵化にも悪影響があるから、造卵数だけによって保護温度を決定することはできない。
地方蚕業試験場が協同で行なった収繭後の保護温度試験においては(1147)、支106号、日110号、日7号、支14号、欧16号、欧18号、支105号その他の品種につき、25℃、23℃、20℃および15℃保護を比較したが、産卵数の最も多かったのは23℃-25℃保護で、これに次ぐのが20℃保護であった。30℃および15℃に保護したものは産卵が著しく少なかった。
23℃-25℃においては不受精卵も最も少なかった。永井(626)の試験においても、20℃、24℃および27℃のうちでは24℃が最もよかった。
農林省蚕糸試験揚が最近の蚕品種について試験した成績をみても(802)、発蛾歩合、正常蚕卵蛾歩合、産卵数などを綜合して、種繭保護の適温は23℃-25℃の範囲にあり、やや委しく云えば
日122号 24℃-25℃ 支115号 25℃
日124号 23℃-24℃ 支122号 23℃-25℃
日125号 〃 支122号(太) 〃
日502号 〃 支124号 〃
支25号 23℃ 支503号 〃
であった、と結論されている。第42表においても、正常卵蛾歩合は蛹を25℃またはその付近の温度に保護した場合に最も高い。。足立ら(13)は産卵量および産卵速度からみて、23℃が最もよかったと報告している。
平山・館石(202)は上蔟後を30℃、24℃、および18℃に保護する3恒温区と、1)化蛹まで、2)化蛹から腹眼着色まで、3)複眼着色から触角着色まで、4)触角着色から発蛾まで、の期間それぞれ毎日、日中(8時から20時まで)30℃、夜間(20時から8時まで)18℃、平均24℃で保護する4変温区とに分けて比較し、第45表に示す結果を得た。
| 試験区 | 春蚕期 | 初秋蚕期 | ||||||
| 調査蛾 数(蛾) |
産卵数 (粒) |
受精卵 歩合(%) |
調査蛾 数(蛾) |
造卵数 (粒) |
産卵数 (粒) |
受精卵 歩合(%) |
産卵歩合 (%) |
|
| 30℃区 24℃区 18℃区 変温1区 変温2区 変温3区 変温4区 |
16 16 14 14 14 10 14 |
494 586 456 571 552 473 585 |
90 98 47 99 95 91 98 |
21 23 21 14 21 21 15 |
484 565 464 550 511 499 570 |
400 556 364 516 483 460 533 |
94 98 62 98 98 97 97 |
82 98 78 93 94 92 93 |
これによると、18℃恒温区が最も悪い。また、平均温度は同じ24℃でも、恒温区に比べて変温区が劣り、変温区相互の間では第3区が悪かった。これは伊与田らが卵巣の発育に対する蛹期保護温度の影響の最も大きい時期としている中期の変温に相当する。造卵数は、伊与田らの結果とは反対に、18℃恒温区が最も少なく、24℃恒温区が最も多く、、第43表の昭和A(黒種)と同様な結果を示した。卵の大小については言及されていないが、1g蟻数が
30℃区 2285頭 変温1区 2562頭
24℃区 2556 〃 2区 2678
18℃区 2895 〃 3区 2683
〃 4区 2632
であったのをみると、卵は、後に述べるように、保護温度の高いほど大きかったものと考えられる。変温区は総べて、24℃恒温区に比べて、1g蟻数が多い(卵が小さい)から、この点からみると、平均温度としては同じでも、変温区は恒温区に比べ、生理的には低目な温度として作用したように思おれる。
1g蟻数の多かったものほど、一般に、蟻蚕の絶食生命時数は短かった。
正常産卵蛾の産卵数は、発蛾したもののうち、産卵数について選抜されたものの産卵数であるから、各区間の差の小さくなるのは当然であるが、第42表において、発蛾歩合や正常産卵蛾歩合に比べ、各区間の差が小さい。特に、後半28℃保護のもの(22℃→28℃)が25℃恒温のものに非常に接近した成績を示しているのが注目される。牛込(1127)が、産卵数は、上蔟後、発蛾までの保護温度68゚F(20℃)または73゚F(22.8℃)のものが、78゚F(25.6℃)または83゚F(28.3℃)のものよりも多いが生殖外器キチン板を透視し得るようになった時期(第36表)以後は、83゚Fに保護しても産卵数が少なくならなかつたと云っているのもこれと同じ傾向を示すものと考えられ、蛹の後期には産卵敬に対する高温の悪影響が減少するものと考えられる。
永井(631)によれば、産絹量の多い系統の蛹は、保護温度21-27℃(湿度75%)の範囲内においては温度の高いほど、湿度63-99%(温度24.5℃)の範囲内では湿度の低いほど産卵歩合(造卵数に対する)が低下したが、これは産絹量の多いものほど吐糸による体水分の減少の多いことに関係があるものと考えられると云う。但し、蛹は裸蛹で保護した。
b 保護温度と卵の大小
蚕の卵は品種によって大きさが異なり、概して、欧州種は支那種および日本種に比べて大きく、日本種の卵は欧州種および支那種に比べて長目である。卵の大きさは、また、蚕の栄養状態によって相違する(Ⅱ2Ac)ほか、同じ蛾の卵でも、初めに産むものは大きく終りのものは小さいが(242,1142)、更に、種繭の保護温度によって相違し、温度の高いほど卵は大きく、1g卵数は少なく、100粒卵重が大きい(第42表)。
木暮・山本(408)によれば、保護温度の高まるに連れて、卵が大きくなると共にその長巾率が小さくなり、卵はまるみを増してくる。宮坂(568)も同様な傾向を報告している。種繭の保護温度が卵の大小に影響する時期は主として化蛹から複眼黒色期までの間で、その後の影響は皆無ではないが少ないと云うから、保護温度が造卵数に影響する時期とほぼ一致している。牛込(1127)は、化蛹前の温度もこれに影響するらしい成績を得ている。
高温保護によって卵の大きくなる理由について、木暮らは、1)包卵被膜細胞の生理作用が増大するのか、2)高温のために発育を中止する卵が生じ、残った卵がその養分を取り込かのか、二つのことが考えられるが、温度の高くなるに連れて卵形の大きくなるのに対して、卵数の変化は温度の上昇と無関係であるから、卵形の大きくなることと造卵数の多少とは無関係であって、上記第2の説明は実際に合わないと結論している。
卵数の変化が温度の上昇と無関係であると云うのは、第43表の昭和A(黒種)の場合を指しているのであるが、これは、卵数を増加させる低温の範囲には、それぞれの場合に応じて限度のあることを示すものとも考えられるから、これだけでは卵数の多少と卵の大小との関係を否定する根拠として不十分なように思われる。卵数が減少してその養分が残りの卵に利用されるのは、氏らの考えているように退化卵からの直接の養分吸収ではなく、発育すべき卵数が制限せられ、配分する養分に余裕が生じ、この分が間接に利用されるものであろう(Ⅰ2Abi)。また卵形に対する温度の影響の最も大きい化蛹から複眼黒色期までの時期は卵巣発育の前期から中期の初めに当り、卵巣は未だ絶対重が小さく、発育も緩徐なときであって、顕著な卵巣の発育は複眼黒色期以後である(第41表)。これらのことと、保護温度が卵の大小に影響する時期と造卵数に影響する時期との一致していることを考え合わせると、卵の大小におよぼす種繭保護温度の影響は2段階に分けて検討すべきもののように思われる。即ち、第1段階として、蛹の初期に温度の直接あるいは直接に近い影響によって、発育すべき卵の数、従って養分配分のプランがきまり、第2段階として、複眼着色期以後の卵の発育によってこのプランが実現されるのであって、卵数の多少と卵の大小とは間接的に関係があるものと考えるべきであろう。
種繭の高温保護によって大きく作られた卵は、催青日数が長く、蟻蚕体重も重いから(第46表)、卵殻ばかりではなく、卵黄の増加していることが明かである。
| 蚕品種 | 保護温度 (℃) |
蟻蚕重量 | 催青日数 | ||
| 調査頭数(頭) | 1頭平均(mg) | 調査蛾数(蛾) | 平均孵化日数(日) | ||
| 昭和A (生種) |
30 25 20 15 |
2,352 2,351 1,017 894 |
0.4252 0.3964 0.3752 0.3365 |
30 - 30 - |
10.2 - 9.5 - |
| 昭和A (黒種) (人工孵化) |
30 20 |
1,725 1,941 |
0.4637 0.4174 |
20 20 |
10.9 10.2 |
直接の実用性は兎も角、造卵数増加の問題の主要項目の一つである卵の数と量との関係の転換(Ⅰ2Abi)が、種繭の保護温度の管理によって一部達成されることは興味がある。
上蔟後の保護温度についてこれまで述べてきた処を綜合すると、全期間恒温の場合には23-24℃、前期と後期との保護温度を区別する場合には、複眼黒色期頃を境にして、前期をやや低目、後期をやや高めにするのがよいと考えられる。産卵が少なく、異常卵の多い傾向のある日122号の蛹を、前期やや低温、後期やや高温に保護して好成績をおさめた例を第47表に示す。但し、不受精卵や催青死卵については、種繭の保護温度以外にも原因の多いことを考慮する必要がある。
| 飼育場所 | 保護温度 | 造卵数 (粒) |
産卵数 (粒) |
異常卵 | ||
| 不受精卵 (粒) |
催青死卵 (粒) |
合計 (粒) |
||||
| A | 1.全期22℃ 2.前期22℃、後期26℃ 3.前期26℃、後期22℃ 4.全期26℃ |
506 520 463 478 |
453 460 424 438 |
22 18 20 23 |
10 8 12 15 |
42 41 38 43 |
| B | 1.全期22℃ 2.前期22℃、後期26℃ 3.前期26℃、後期22℃ 4.全期26℃ |
654 650 590 582 |
589 597 542 540 |
21 22 25 29 |
12 12 15 16 |
51 50 54 50 |
| C | 1.全期22℃ 2.前期22℃、後期26℃ 3.前期26℃、後期22℃ 4.全期26℃ |
510 502 432 440 |
443 439 418 425 |
18 15 20 25 |
9 10 13 12 |
45 41 39 44 |
c 保護温度と卵の越年性
種繭の保護温度と卵の越年性との関係については、渡辺(1139,1141)によって、高温保護は不越年性卵を多くすることが報告されていたが、最近、休眠ホルモンを注射した不越年卵性蛹(蛾になってから不越年性卵を産む蛹の意味である。以下これに準ずる。)は、注射後を高温におくよりも、15℃
に5日間ぐらい保護した方が越年性卵を多く産むことがわかって(165,169)、ホルモン作用の面からも証明された。
しかし、この渡辺の報告は、二化性または四化性種の卵を20℃前後の中間温度で催青した場合の蚕を飼育して得た蛹の保護温度について述べたもので、普通のように二化性種を高温(25℃)で催青した場合には、上蔟後の保護温度がかなり高いときにも不越年性卵は出にくい(第48表)。
| 催青温度 | 4令期以後の保護温度 | 産卵結果 |
| 25℃以上 | 飼育中25℃以上、上蔟後30℃ 〃 17℃ 、 〃 20℃ |
越年卵 〃 |
| 15℃以下 | 飼育中25℃以上、上蔟後30℃ 〃 17℃ 、 〃 20℃ |
不越年卵 〃 |
| 20℃前後 | 飼育中25℃以上、上蔟後30℃ 〃 17℃ 、 〃 20℃ |
越年卵と不越年卵(不越年多し) 〃 (越年多し) |
この表には、4令期以後の飼育温度17℃の場合も示してあるが、これでわかるように、二化性種を25℃で催青した場合には、普通には、飼育温度は稚蚕期、壮蚕期とも化性に影響することが少ない。
渡辺(1141)は、産卵の越年現象を、卵の発育を一時的に抑制するある物質の存在によって起こるものと仮定し、この物質を抑制質(inhibitory
substance)と名什けた。この説明によると、二化性種を高温で催青した場合には、胚の体中に多量の抑制質が生成し、これが蚕児、蛹を径て次代の卵に伝わり、その卵を越年させるが、低温で催青すると抑制質が生成されないため、次代の卵が不越年性になるのである。このように催青温度が高温または低温であると、卵の越年性または不越年性は安定しており、上蔟後の温度によって影響を受けないが(第48表)、中間温度で催青すると、抑制質の生成される胚と生成されない胚とが生じ、生成されたものにおいても、抑制質の量は個体間に変異がある。抑制質の少い蚕は、4令期以後、卵巣内で卵の分化の始まる頃からの保護温度の影響により、温度が比較的低いときには、少量の抑制質によってもその卵巣内の卵を越年化させることができるが、温度が高いと抑制質の作用が発現し得ず、卵は不越年性になると云うのである。
上蔟後の保護温度が低いときに再出卵の多いことも、同様に抑制質の仮定によって説明されている(1143,1144)。ここに再出卵と云うのは、漿膜が着色して越年性卵の外観を呈する卵でありながら、産卵後2週間前後で孵化し、1蛾の卵の中に越年性卵と混在しているもののことである。
中間温度で催青した抑制質の少ない蚕を、上蔟後、中間温度で保護すると、抑制質の作用がある程度働くために、完全な不越年性にはならずに産み出されるが、越年性が不安定な状態にあるため、産卵後の保護温度が高いと再出卵になる。これに対して、上蔟後の保護温度が高温であると、抑制質の少ない蛹の卵は不越性卵になることが多く、不安定な状態で産み出されるものが少ない。従って、再出卵も少ない。即ち抑制質の少ない卵が、保護温度の相違によってあるいは再出卵になり、あるいは不越年性卵になるのであるから、不越年卵蛾区と再出卵蛾区との合計は、上蔟後の保護温度が高くても中間温度でも、殆ど変わりがないことになる(第49表)。高温催青を行なえば、この場合の再出卵は殆どあるいは全く生じない。
| 試験 | 上蔟後発蛾 までの温度(℃) |
総蛾区数 (蛾) |
生種蛾区 数(蛾) |
再出卵蛾 区数(蛾) |
(生種蛾区+再出卵蛾区)/総蛾区数×100 (%) |
| A | 30 20 |
144 227 |
53 15 |
20 115 |
50 57 |
| B | 30 20 |
122 216 |
67 71 |
7 49 |
60 55 |
抑制質による越年性の決定は、産卵後、低温に保護すると数日中に行なわれるから、産卵後12時間以内に15℃に移して3日間保護すれば、その後は高温に戻しても再出卵は殆ど発現せず、15℃に5日間おけば全く発現しないのが普通であると云う(Ⅴ2B)。
これに対し、木暮(406,409)は、蛹の保護温度が高いと着色卵中の不越年卵(再出卵)歩合は減少し、着色卵が再出卵になるか否かは催青中の温度および光線とは無関係であると考えた(Ⅴ2A)。室賀(610,616)は、木暮と同様に、催青温度と着色卵中の再出卵歩合との間には関係のないことを認めたが、上蔟後、発蛾までの温度が高いと、着色卵蛾歩合が減少して不着色卵蛾歩合の増加すること、および不着色卵中の越年性卵歩合が高まると云う結果を挙げて、蛹の高温保護が再出卵を少なくするようにみえるのは、抑制質の少ない越年性の不安定な着色卵蛾が不着色卵蛾に変わり、残った着色卵蛾の産む卵には再出卵が少ないためであって、本質的に越年性卵を増加させるのではないと云い、越年性と着色性とは共に催青中に決定されて、互に密接な関係があるものと考えている。
河合(355)は、大造系統の材料を用い、25℃で催青し、蛹期を20℃と30℃とに分けて保護したが、後者に再出卵が多く、3・ヒドロキシキヌレニンの卵内移行量からみても越年性の不完全な卵であった。これは、渡辺の場合と異なり、25℃催青であるから、休眠ホルモンの分泌は十分であり、蛹期間を20℃で保護すればその越年性が一層完全になるのに対し、30℃保護においては、高温のために越年性物質(抑制質)および着色物質の卵内への移行が少なく、そのために不完全な越年性卵が生じ、再出卵になったものと解釈している。
一般に再出卵と呼ぶものの中には、上記のものとは性質の異なったものが合まれているが、これについては後に述べる(Ⅶ1A)。
ここで注意しなければならないのは、15℃に保護すると越年性が決定するとは云っても、長く15℃におき、高温(例えば25℃)に戻さなければ、越年性はいつまでも完成しないことである。
155℃に保護し続けると、60日たっても、その越年性(活性化の難易および呼吸の青酸阻害からみた)は25℃保護の産卵後2日目ぐらいのままに止まり、越年性卵ではあるが非常に活性化し易く(785)、安全な蚕種保護を行なうことができない。大野(767)によれば、15℃に15日間おいて25℃に戻したものは、翌年の孵化歩合が著しく悪かった。 これは越年性が不完全で、早く活性化する卵ができるためである。
10日までは異常がなかったと云うが、必要な日数の後には早く25℃に移して休眠の完成をはかる必要がある。
牛込(1127)は、保護温度を上蔟後の時期別に変化させて行なった試験の結果から、化蛹までを83゚F(28.3℃)に保護すると、この期間を68゚F(20℃)に保護した場合に比べて不越年性卵の多い傾向があると云っているが(第50表)、これは休眠ホルモンの作用に温度の影響する時期とは少しずれている(Ⅴ2C)。
| 蚕品種 | 保護温度(゚F) | 産卵蛾数 (蛾) |
不越年卵混産 蛾歩合(%) |
不越年卵蛾 歩合(%) |
不越年卵 歩合(%) |
||
| 前期 | 中期 | 後期 | |||||
| 日110号 | 68 73 78 83 68 68 83 68 83 83 |
68 73 78 83 68 83 68 83 68 83 |
68 73 78 83 83 68 68 83 83 68 |
173 176 244 155 179 208 198 199 211 222 |
0 0.6 0 0.7 0 0 1.1 0 0 1.0 |
0 0 0.4 3.3 0.6 1.5 1.1 1.6 3.8 0.5 |
0 0.1 1.0 4.0 0 1.2 1.0 0.6 4.0 0.6 |
| 支102号 | 68 73 78 83 68 68 83 68 83 83 |
68 73 78 83 68 83 68 83 68 83 |
68 73 78 83 83 68 68 83 83 68 |
126 147 151 186 105 128 112 105 110 162 |
0 3.7 0 1.4 1.0 0.9 9.4 1.0 0 0.6 |
2.7 4.4 2.7 8.5 2.0 2.7 0.9 0 6.4 7.5 |
1.7 4.7 2.8 8.4 1.9 2.5 5.5 0.5 6.6 5.1 |
d 保護温度と孵化の良否
i 越年種 高梨・松尾(957)が支109号を用い、上蔟から発蛾までの間を、28℃、25℃および20℃の3区に分けて保護し、採種後は何れも25℃に保護しておき3、10、20、30、50、70、90、120、150、180および210日後に5℃に移し、所定の日数毎に出庫して25℃で催青した結果の一部を第51表に示す。
これによると、90%以上の孵化歩合の続いた冷蔵期間は、上蔟後を25℃に保護したものが最も長く、20℃保護がこれに次ぎ、28℃保護が最も短かかった。また各冷蔵日数毎の孵化歩合も一般に25℃区が最もよく、これに続いて20℃区、28℃区の順位であった。表に示したのは総孵化歩合であるが、実用孵化歩合の順位もこれと同じであった。高梨らは、このような結果の得られた理由には触れていないが、蚕種保護上の重要な問題を含んでいる。
| 冷蔵日数 (日) |
高温日数 | |||||||||||||||||
| 3日 | 10日 | 30日 | 90日 | 180日 | 210日 | |||||||||||||
| 上蔟後の温度 | ||||||||||||||||||
| 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | |
| 30 50 60 70 80 90 100 120 140 160 180 200 220 240 260 280 300 325 |
4% 75 96 97 97 97 96 98 97 96 98 93 91 78 56 30 7 - |
8% 86 92 92 95 91 93 96 97 97 97 90 77 39 13 2 6 0 |
% 61 86 86 91 89 88 90 90 90 91 83 83 65 26 8 2 - |
0% 20 70 94 96 96 93 94 97 97 96 96 92 88 73 44 22 7 |
0% 23 89 91 93 92 93 95 97 97 93 94 91 81 54 32 9 1 |
0% 27 44 76 89 94 91 91 93 97 93 86 89 67 44 17 3 1 |
0% 21 62 94 95 96 96 97 97 96 97 96 95 90 82 69 48 11 |
% 25 62 91 95 95 96 94 95 98 97 96 93 85 68 34 16 5 |
0% 14 43 75 93 93 92 93 92 92 91 93 88 88 75 56 34 19 |
0% 4 16 44 73 84 91 95 95 96 81 89 83 73 46 37 18 7 |
0% 4 14 51 73 86 90 95 92 92 92 89 71 55 38 23 14 3 |
0% 1 17 51 68 83 91 93 94 93 90 82 85 67 59 43 23 8 |
0% - 54 72 80 85 88 92 89 80 69 59 - - - - - - |
0% - 39 57 65 74 78 81 77 70 57 43 - - - - - - |
0% - 48 63 84 80 86 88 88 78 55 59 - - - - - - |
0% - 78 86 86 82 84 76 72 61 46 20 - - - - - - |
0% - 61 76 63 51 53 54 39 35 21 12 - - - - - - |
0% - 75 77 78 75 74 69 62 45 34 19 - - - - - - |
まず、90%以上の孵化歩合を示し初めたときの孵化歩合および冷蔵日数から考えると、25℃区の活性化が20℃区よりも早いと判断されるのであるが、高温期間50日頃までは、20℃区は25℃区に比べて、活性化初期(孵化の始まる最短冷蔵日数)の孵化歩合が僅かながら殆ど常に高く、冷蔵期間が長くなって孵化歩合の低下し初める状態をみると20℃区が25℃区よりも常に早い。これは、20℃区には活性化の特別に早い卵が少数含まれていることを示すような成績である。
28℃区は活性化初期の孵化歩合が3区の中で最も低く、また孵化歩合が90%に達するまでの冷蔵日数が最も長い。これは活性化のおくれていることを示す成績のように思われる。
20℃区と28℃区とを比較すると、高温期間の短い間は、極端に冷蔵期間の長い場合を除けば、常に20℃区の孵化歩合が高いが、高温期間150日頃を境に逆転して20℃区の孵化歩合が低くなる。この時期においても、孵化歩合の最も高いのは25℃区であるから、20℃区は、25℃区および28℃区に比べて、高温期間の持続によって孵化機能を失い易い卵を合んでいるもののようである。これが、20℃区に抵抗力の弱い卵が合まれているためか否かは、与えられた数字だけからは不明であるが、20℃区には、25℃区および28℃区に比べて生理的に不安定な状態の卵の多いことは確かであろう。
20℃区に活性化の早い卵のできることは、あるいは、前項で述べたように、抑制質の少ない卵が不安定な越年性のまま残っているためであると考えら)れないこともないが、28℃区に活性化のおそい卵のできることは、休眠の成立についての渡辺の説明だけでは理解の困難な現象のように思われる。
滝沢・勝野(1005)は、上蔟後の保護温度25℃、20℃および15℃について、上と同様な方法で比較を行ない、25℃区の孵化が最もよく、15℃区の孵化の最も劣ることを観察し、また、蛹期間を前、中、後の3期に分け、25℃と15℃とを組合わせて保護した結果では、前、中期を25℃で保護したものの孵化が最もよく、この時期を15℃で保護したものは最も悪かったと述べているが、前、中、後期の分け方は不明である。
以上は人工越冬によって卵を活性化させて比較した孵化試験であるが、普通に越年種の保護をした場合にも、上蔟以後の温度によって翌春の孵化状態に相違ができる。一例として、昭和37年度関東地区蚕業試験場協力試験の結果を挙げる(印刷発表されていない)。
試験時期は春蚕期で、分場農家が上蔟後何時頃まで補温を行なうかを想定して、次ぎの3区を設けた。
A 上蔟後まる3日間を25℃目標で保護し、以後収繭まで自然温度で保護
B 上蔟後、収繭までを通して25℃目標で保護
C 上蔟後まる6日間を25℃目標で保護し、以後収繭までの短期間を自然温度で保護
収繭後は、各場所の慣習により普通の保護、採種を行ない、交雑種を作った。蚕種は普通に保護した後、12月上-中旬に洗落してバラ種とし、冬期間を通じて毎月1回孵化調査を行なった。
結果の中から群馬県蚕業試験場の成績を示すと第52、53、54表の通りで、産卵状態はA区が最も悪く、B、C両区の間の差は明かでない。
A区は活性化初期の孵化歩合がやや高かったが、最高孵化歩合を示す時期は3区間に殆ど差がなかった。洗落し後を5℃で通したため春蚕催青期には3区とも孵化歩合がかなり低下していたが、特にA区の低下が大きかった。これらの結果は何れも、人工越冬によって試験した第51表の傾向と一致している。
| 蚕品種 | A区 | B区 | C区 | |||||
| 上蔟後 3日間 |
以後収繭 まで |
収繭から 発蛾まで |
上蔟から 収繭まで |
収繭から 発蛾まで |
上蔟後 6日間 |
以後収繭 まで |
収繭から 発蛾まで |
|
| 日124号 支122号(太) |
25.0℃ 25.0 |
19.7℃ 20.0 |
25.7℃ 25.7 |
25.0℃ 25.0 |
25.6℃ 25.6 |
25.0℃ 25.0 |
21.0℃ 21.0 |
25.7℃ 25.6 |
| 蚕品種 | 区別 | 正常蚕卵 蛾 (%) |
少数蚕卵 蛾 (%) |
不受精 卵蛾(%) |
再出卵蛾 (%) |
無蚕卵蛾 (%) |
| 日124号×支122号(太) | A B C |
70 84 81 |
8 9 9 |
13 6 6 |
0 0 0 |
8 0 3 |
| 支122号(太)×日124号 | A B C |
25 84 65 |
15 4 4 |
57 10 28 |
0 0 1 |
2 0 0 |
| 蚕品種 | 区別 | 洗落し 翌日 |
出庫月日 | 春蚕催青 の時 |
|||
| 12月25日 | 1月20日 | 2月20日 | 3月20日 | ||||
| 日124号 × 支122号(太) |
A B C |
2% 0 1 |
62% 40 52 |
95% 95 94 |
95% 93 94 |
96% 94 95 |
66% 67 73 |
| 支122号(太) × 日124号 |
A B C |
3 0 2 |
56 38 40 |
96 92 93 |
95 97 97 |
96 96 97 |
80 92 93 |
ii 人工孵化種 渡辺(1149)は、上蔟から蛾発までの保護温度20℃-30℃の範囲内においては、保護温度の高いほど、有効な孵化歩合の得られる即浸浸漬時間の巾が広いと報告している(第55表)。しかし、足立ら(9,10,11)の現行品種についての試験成績では、品種によっても相違があり、このような明瞭な関係を認め得なかった。
| 浸漬時間 | 保護温度 | |||||
| 30℃ | 25℃ | 20℃ | ||||
| 総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
|
| 30秒 1分 2 3 5 10 12 15 20 25 30 35 40 |
26 90 95 97 93 96 90 93 91 96 68 19 4 |
19 78 84 92 89 91 89 92 90 90 67 17 3 |
40 97 98 98 98 95 98 98 94 75 81 20 0 |
37 96 98 97 98 94 98 97 91 72 70 14 0 |
81 96 97 96 96 97 91 88 79 31 33 12 0 |
74 81 97 96 96 96 86 63 61 20 20 6 0 |
短期冷蔵浸酸についての渡辺の成績は第56表の通りで、即時浸酸の場合と同様、90%以上の孵化歩合の得られる浸酸時間の巾は上蔟後の保護温度の高い場合に広かった。
渡辺は、蚕の化性は上蔟から発蛾までの期間において保護温度の影響をうけるので(1139)、人工孵化の成績も種繭保護温度の高低によって相違するのではないかと考えて、この試験を行なったと述べているが、これらの結果と化性との関係については言及していない。
第55表をみると、90%以上の孵化歩合を示す浸漬時間の巾はなるほど30℃区が最も広いが、最も短い浸酸時間での孵化歩合をみると、保護温度の低いほど高いから、低温区において浸漬時間の巾の狭いのは、塩酸刺戟に対する感受性が強く、浸酸時間が長いと被害を受けて孵化し得ないのであって、低温保護のために越年性が安定して浸酸が効きにくいのではない。上蔟後の保護温度が低いと卵が小さく(Ⅲ3Bb)、卵が小さいと塩酸刺戟に対する感受性の強い(Ⅷ1A)ことがこの原因ではないかとも考えられるが、明かにされていない。
| 浸漬時間 (分・秒) |
保護温度 | |||
| 25℃ | 20℃ | |||
| 総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
|
| 1・0 1・30 2・0 2・30 3・0 5・0 6・0 7・0 8・0 10・0 12・0 15・0 |
93 91 96 96 98 97 95 72 68 2 2 0 |
87 85 92 93 95 90 79 70 67 2 2 0 |
73 95 96 96 96 93 73 50 21 4 1 0 |
63 87 94 94 95 92 71 46 16 2 0 0 |
沓掛(475)は、上蔟から発蛾までを25℃と15℃ とで保護した種繭からの卵の即時浸酸について、第55表と同様な成績を示しているが、上蔟から化蛹までを前期、化蛹から複眼着色までを中期、以後発蛾までを後期として、25℃と15℃とを組合わせて時期別に保護温度を違えると、90%以上の孵化歩合を示す浸漬時間の巾に最も大きく影響するのは中期の温度であると云う。この時期の保護温度が卵の大小や休眠に影響することは既に述べた。
冷蔵浸酸は、産卵後約2日間高温で保護して越年性を進めた卵を冷蔵後に処理するのであるから、即時浸酸の場合と違い、越年性が成績に影響する筈である。第56表において、卵の小さい筈の20℃区の孵化歩合が、最短浸酸時間では最も低く、最高孵化歩合を示す浸酸時間を超えると、他の2回に比べて早く低下しているのは、越年性と卵の大きさとの両者が共に関係していることを示すものであろう。
小野寺(715)が、支110号の同蛾区の熟蚕を3分して、上蔟後、発蛾までをそれぞれ30℃、25℃および20℃に保護したものから採種し、同一母蛾の卵を分割供試して行なった蚕種の抵抗力試験によると、即時浸酸後3日目から40℃、湿度75%に、24時間接触した場合の抵抗力指数(対照区の孵化歩合を100とする)は、30℃区(供試蛾数37蛾)64.8±1.9、25℃区(23蛾)48.5±2.1、20℃区(37蛾)44.2±1.5で、上蔟後の保護温度の高いほど抵抗力が強く、特に30℃区の指数は著しく大きかった。
また、浸酸後3日目の卵を蒸溜水に24時間浸漬した場合には、30℃区(37蛾)70.5±2.5、25℃区(26蛾)44.2±2.9、20℃区(37蛾)36.4±2.8、浸酸後2日目の卵を無水アルコールに50分間浸漬した場合には、30℃区(17蛾)28.8±3.0、25℃区(22蛾)2.7±1.0、20℃区(18蛾)1.1±0.3、4日目の無水アルコール浸酸30分間の成績も同様な傾向を示し、常に保護温度の高い区において抵抗力の大きいことが注目される。
この原因には言及されてていないが、上記渡辺の浸酸試験の場合と共通の原因が関与しているように思われる。
なお小野寺は、点青期における12時間浸水についても同様のことをみている。
4 発蛾調節のための蛹の保護
A 温度管理
発蛾調節のための蛹の保護温度は、蛹期間を短縮または延長させる程度の温度と発育を殆ど停止させる温度(冷蔵)とに区別される。
蛹期間の短縮または延長は、蛹の発育階程に基ずいて計画的に行なうもので、例えば、第38表において、25℃で保護してきた日122号を複眼着色期に28℃に移すと、発蛾までの日数を2日間短縮でき、同じく25℃で保護してきた支115号を複眼黒色期に22℃に移せば、発蛾までの日数を1日間延長することができる。
30℃または15℃を使えば、第41表のように、更に大巾な短縮、延長を行なうことができるが、28℃以上の高温、20℃以下の低温保護は、前節で述べた各種の成績からみて行なうべきではない。なお、蛹の外形に基ずいて温度管理を行なっても、判定の誤差や品種の相違などのために予定通りの調節ができず、次ぎに述べる冷蔵法によって再調整する必要のあることが多い。
発育停止に近い温度による冷蔵法の施行は、その時期、期間および温度について、前法以上に注意をする必要かおる。冷蔵時期とその温度については多くの報告があり、不一致の点も少なくないが、一般に適期とされているのは複眼の着色し初めた化蛹3-5日目頃と発蛾前日であり、温度は2.5℃-7.5℃が最も多く用いられている。
藤本(100)の行なった雌蛹の冷蔵試験によれば、25℃で保護中の蛹を2.5℃に7日間冷蔵し、その後は再び25℃に戻して調べた処、化蛹第1日目および2日目冷蔵のものは冷蔵日数以上に蛹期間が延び、それぞれ18.8±0.04日および18.9±0.04日(無冷蔵は11.4±0.03日)となったが、その後化蛹10日目(11日目に発蛾)までの間に冷蔵したものでは、冷蔵期間以上の発蛾のおくれは起らなかった。発蛾歩合は全体的に低下するが、特に化蛹第1日目および2日目に冷蔵したものにおいて最も低くかった。造卵数は化蛹1-7日目、産卵歩合は1-6日目および9-10日目、不受精卵歩合は1-5日目に冷蔵したものがそれぞれ無冷蔵と大差なく、その他の時期の冷蔵においては総べて無冷蔵より劣った。従って、これらに共通してよかった冷蔵時期は化蛹3日目である。
化蛹3日目の冷蔵につき、2.5℃、5℃、7.5℃、10℃、各7日間の冷蔵を比較すると、発蛾歩合は全般に無冷蔵に劣り、試験した範囲の温度では、その高低と発蛾歩合との間に関係が認められなかった。造卵数は、2.5℃-7.5℃冷蔵、産卵歩合は2.5℃冷蔵、不受精卵歩合は2.5℃-5.0℃冷蔵がそれぞれ無冷蔵と差がなく、共通的によかったのは2.5℃であった。
化蛹3日目冷蔵の場合の2.5℃および5℃冷蔵では、無冷蔵(11.2±0.03日)に比べて冷蔵日数(7日間)だけ蛹期間が長くなったが、7.5℃冷蔵では18.0±0.05日、10℃冷蔵では17.5±0.03日で、何れも、冷蔵日数を加えただけは蛹期間が延びなかった。これは、これらの温度においては、冷蔵中に蛹の発育することを示すものである。藤本の実験結果を綜合すると、蛹期間が10日の場合の化蛹3-5日目(上蔟7日目)に2.5℃に冷蔵すれば、発蛾は多少不斉になるが、その他の点では、最長5日間までは無冷蔵と変りがないと云うことになる。但し、品種は支那二化性S2である。化蛹3日目の蛹の外形は記載していない。
牛込(1127)の成績をみても、化蛹3日目(複眼の着色し初めた時期)の冷蔵が最もよく、発蛾前日がこれに次ぎ、化蛹7日目(生殖外器キチン板が黒色にみえ初める時期)の冷蔵は最も悪い。冷蔵温度は41゚F (5℃)がよかった。
大宮(739)が日122号および支122号(太)の蛹を30℃に3日間、5℃に7日間、または0℃に7日間おいて造卵数を調べた成績では、複眼着色期に処理したものが、腹眼黒色期に処理したものよりも常に造卵数が多かった。産卵歩合には処理時期による差がなかった。
戸谷(1069)によれば、化蛹5日目の蛹(日124号、蛹期間12日)を5℃に10日間冷蔵し、発蛾後解剖して卵管を調べると、卵管下部に卵殻形成の不完全な卵がみられる。化蛹8日目の冷蔵ではこの異常卵の分布する部位が卵管の中部となり、更に冷蔵時期がおそいとこれが卵管の上部に分布するようになる。産卵させてみると、これらの異常卵は早期潰れ卵になるが、冷蔵時期がおそいほどこれの産み出される順位もおくれ、卵管内で認めた異常卵の分布関係とほぼ一致した。異常卵は卵殻に穴のあるもの、または卵殻が内層のみで中、外層を欠くものなどであったが、切片にして調べた結果、異常卵になるのは包卵被膜細胞が完成しかけた時期に冷蔵した卵であることがわかった。第36表について見ても、化蛹5-6日目の蛹の卵管下部は丁度この時期の卵で占められており、蛹令の進むに連れてその部位が上昇する。この実験において(化蛹7日目の冷蔵で潰れ卵が最も多く、3日目の冷蔵では潰れ卵の発現しなかったことは上に述べた冷蔵試験の結果とも一致している。10日間にも亘る冷蔵は実際には行なうべきものではないが、冷蔵は蛹の発育階程を考えて行なう必要のあることを示す好例である。
横山・丸茂(1200)は日122号(細)および支115号を用い、上蔟後25℃、湿度75%目標で保護した種繭を、複眼の着色し初めた頃と発蛾前日とに5℃、湿度80%目標で3日および7日間冷蔵したが、複眼着色期に冷蔵し、出庫時に10℃および20℃各6時間の中間温度を通したものが発蛾歩合、産卵数、孵化歩合、催青死卵、不受精卵などの調査において最もよく、冷蔵3日と7日との間には余り差がなかったと云う。発蛾前日の冷蔵は冷蔵日数の長くなるほど成績が悪かった。
以上の結果を通覧すると、蛹の冷蔵は複眼の着色し初める頃に行なうのが最もよいが、発蛾調節のための冷蔵は初めから予定して行なうものではなく、応急措置であるから、長い期間の冷蔵では劣るとしても、作業上からは発蛾前日に行なうことが多い。
発蛾前日の冷蔵は3日間でも採種成績に影響するが、冷蔵の影響は、裸蛹と繭殻中に収めたものとでも相違する(第57、58表)。
無冷蔵の場合でも、裸蛹にしたものは、切り出し時期の早いほど産卵数が少なく、不受精卵の多いことが古くから云われている(530)。 裸蛹にすると健蛹数が確認できること、発育階程を監視し易いことなどの便宜のある代りに、温湿度などの環境変化の影響を直接に受け易いこと、外傷や病気の感染を受け易いこと、カツオブシムシなどの虫害も多いことなどの欠点を伴なっている。死籠りの多い場合などには切り出し作業に注意しないと、病気を蔓延させることがある。蛹期に膿病ウイルスを接種すると(複眼赤色のとき)、品種によってかなりの遅速はあるが、早いものは82時間(28℃の場合)で発病すると云う(21)。蛹の外皮の最外層は体内水分の発散防止に役立っているから、アルミナの様なもので体表面をこすると体水分の発散が多くなって、正常に発蛾し得ないもののできること、この水分発散防止には脂質の存在が関係しているので、キシロールなどで蛹体面を拭っても水分の発散が多くなり、蛹重の減少すること、などが知られているが(279,283,284)、また外皮の脂質には硬化病に対する抗菌性もあると云うから(410)、切り出した蛹の取扱いは丁寧にする必要かおる。永井(628)は雌蛹の出血が造卵数および産卵数に影響することを報告している。雌蛹の出血は浸酸成績にも影響する(Ⅷ1A、Ⅶ2Aa)。
| 蚕品種 | 試験区 | 発蛾歩合(%) | 産卵状態(蛾の百分率)(%) | ||||||||
| 冷蔵 | 蛹の状態 | ♀ | ♂ | 不産卵 | 少数産卵 | 不受精卵 | 不越年卵 | 累積卵 | その他 | 正常卵 | |
| 日124号 | 無冷蔵 | 切繭中 | 98 | 100 | 4 | 15 | 11 | 0 | 0 | 1 | 67 |
| 3日間 | 切繭中 裸 蛹 鋸屑中 |
92 90 91 |
100 98 99 |
4 2 9 |
20 20 14 |
9 15 11 |
0 0 0 |
1 1 1 |
1 1 1 |
63 60 62 |
|
| 7日間 | 切繭中 裸 蛹 鋸屑中 |
87 79 84 |
100 91 100 |
5 11 7 |
14 20 15 |
14 19 13 |
0 0 0 |
1 0 1 |
1 1 1 |
62 48 61 |
|
| 支124号 | 無冷蔵 | 切繭中 | 93 | 97 | 4 | 8 | 4 | 0 | 1 | 1 | 80 |
| 3日間 | 切繭中 裸 蛹 鋸屑中 |
91 90 96 |
100 95 97 |
5 7 6 |
7 5 8 |
4 5 4 |
0 0 0 |
0 1 1 |
1 1 1 |
81 78 79 |
|
| 7日間 | 切繭中 裸 蛹 鋸屑中 |
83 77 81 |
91 90 91 |
8 7 7 |
8 5 7 |
6 18 14 |
0 0 0 |
2 2 1 |
2 2 1 |
72 63 67 |
|
| 蚕品種 | 試験区 | 正常産卵蛾1蛾当り | 雌1蛾当り | 1g卵数 | |||||
| 冷蔵 | 蛹の状態 | 総産卵数(粒) | 正常卵(%) | 死卵(%) | 不受精卵(%) | 粗卵量(g) | 精選卵量(g) | ||
| 日124号 | 無冷蔵 | 切繭中 | 539 | 95 | 0 | 4 | 0.218 | 0.179 | 1,649 |
| 3日間 | 切繭中 裸 蛹 鋸屑中 |
503 457 526 |
94 96 94 |
0 0 0 |
5 3 5 |
0.198 0.197 0.200 |
0.159 0.167 0.161 |
1,630 1,638 1,615 |
|
| 7日間 | 切繭中 裸 蛹 鋸屑中 |
490 446 472 |
91 88 92 |
0 0 0 |
7 11 7 |
0.201 0.150 0.187 |
0.167 0.111 0.159 |
1,613 1,610 1,613 |
|
| 支124号 | 無冷蔵 | 切繭中 | 526 | 92 | 0 | 7 | 0.254 | 0.237 | 1,773 |
| 3日間 | 切繭中 裸 蛹 鋸屑中 |
509 493 501 |
91 88 91 |
1 2 0 |
6 9 8 |
0.215 0.201 0.205 |
0.202 0.185 0.192 |
1,770 1,780 1,766 |
|
| 7日間 | 切繭中 裸 蛹 鋸屑中 |
477 472 482 |
82 82 82 |
0 0 0 |
16 17 17 |
0.195 0.187 0.189 |
0.172 0.160 0.162 |
1,790 1,771 1,780 |
|
第57、58表の場合は何時頃切り出した蛹であるのか明かでないが、鋸屑に埋めて冷蔵したものの成績が切繭で冷蔵したものと殆ど差異のないのをみると、中間温度を通したものの成績がよかったと云う報告(1200)と合わせ考えて、急激な温度変化によって生理的な障害を蒙ったことが裸蛹の成績を悪くした主な原因と考えられる。蛹に-5℃、24時間または48時間の低温衝撃を与えた場合にも繭層の保護作用が認められると云う。但し、繭腔内温度の測定結果をみると、外温の変化に対する繭層の防護作用は極めて小さい(1193)から、気温ではなく容器の熱伝導が影響しているように思われる。この点を工夫すれば裸蛹冷蔵の被害を軽威することができるかも知れない。
裸蛹で保護するとカツオブシムシの被害を蒙り易いが、新らしい杉の本の鋸屑を日乾したものに蛹を埋めておくと被害が軽減できると云われ、これの手に入り易い地方では実際に用いられている。これは杉鋸屑の匂いがカツオブシムシの忌避剤として作用するのであろうと考えられる。裸蛹を木箔に収容し、杉の精油を加えた籾殻(籾殻100gに対し杉の粗精油0.25-4.00ccの割合に混合)に埋め、蓋をして保護し、発蛾歩合および交尾、産卵を調べたが、蛹に対する杉油の害は認められなかった(875)。杉鋸屑は精油1%内外を含むに過ぎないと考えられるから、カツオブシムシに対する効果は兎も角、蛹の保護材料に用いて害作用はないものと思われる。
B 光線管理
蚕の蛾は、毎朝明るくなると発蛾し初め、数時間内にその日の分は出てしまうのが普通であるが、品種によっては、発蛾がだらついたり、または朝非常に早く発蛾して暴れるために、雄蛾がいたんで採種上の障害になることがある。支115号、支124号などは後者の例であるが、これらの品種においても、光線管理を適当に行なえば発蛾時刻を調節することができる。
第59表は支115号の成績であるが、支124号についても全く同様な結果が得られている。但し、表において8時の発蛾となっているものは27時から8時までの間(8時に近い)の発蛾である。これは、この品種においては、暗中に保護しても12時間までは発蛾を抑えることができず(抑えられるのは10時間以内)、暗中で発蛾し初めるためであって、明暗交替を4時間毎にすると、大部分が明にしたときから発蛾し初める。また、明暗交替の時刻を8時と20時とにとったため、この表では発蛾が8-12時に集中しているが、明暗交替の時刻を希望の時刻、例えば4時と16時とにとれば、発蛾を4-6時に集中させることができる。
明暗交替を始めるのは初発蛾の出る1日前からでよく、収繭後このときまでの明暗は関係がない。明るさは種繭を収容した箔の上面で5lux以上ならば大体よいが、40lux以上あれば申分ない。ただし、暗くしておく間は完全な暗黒が望ましく、5lux程度のうす暗がりでは暗としての効果がない。
蛹の眼、頭頂白色部および腹部に即乾性黒色エナメルを塗ったり、ゴム板で遮蔽したりして試験をしてみると、程度の差はあるが、体表の大部分が光に対して感受性を持っている(434)。しかし、主要な光感受部位は頭胸部にあると推定される(423)。眼と頭頂白色部との何れの光感受性が大きいかはなお不明である(424)。
| 光線管理 | 性別 | 6月13日 | 6月14日 | 6月15日 | 6月16日 | ||||||||||||||||||||
| 4 | 8 | 12 | 16 | 20 | 24 | 4 | 8 | 12 | 16 | 20 | 24 | 4 | 8 | 12 | 16 | 20 | 24 | 4 | 8 | 12 | 16 | 20 | 24 | ||
| 収繭(上蔟7日 目)以後発蛾 終了まで恒明 |
♀ ♂ 計 |
3 1 4 |
5 17 22 |
9 15 24 |
9 6 15 |
8 4 12 |
7 9 16 |
13 12 25 |
21 25 46 |
5 2 7 |
8 6 14 |
3 1 4 |
3 3 |
4 2 6 |
2 2 |
1 1 |
2 2 |
3 3 |
|||||||
| 収繭以後 発蛾終了 まで恒明 |
♀ ♂ 計 |
3 3 |
4 2 6 |
3 8 11 |
4 4 |
2 13 15 |
13 25 38 |
16 11 27 |
12 12 24 |
3 7 10 |
3 3 6 |
5 9 14 |
46 9 55 |
13 13 |
2 2 |
1 1 |
2 1 3 |
1 1 2 |
1 1 |
||||||
| 収繭後恒明、 初発蛾当日 から明暗交替 |
♀ ♂ 計 |
3 2 5 |
11 11 |
3 3 6 |
2 6 8 |
20 20 |
66 58 124 |
24 3 27 |
15 10 25 |
10 10 |
4 1 5 |
||||||||||||||
| 収繭後恒明、 初発蛾1日前 から明暗交替 |
♀ ♂ 計 |
5 31 36 |
8 2 10 |
33 57 90 |
28 5 33 |
1 1 |
1 1 |
19 26 45 |
12 2 14 |
1 1 2 |
|||||||||||||||
| 収繭後恒暗、 初発蛾1日前 から明暗交替 |
♀ ♂ 計 |
2 9 11 |
2 2 |
11 67 78 |
36 1 37 |
1 1 |
1 1 |
42 21 63 |
29 1 30 |
4 3 7 |
2 2 |
||||||||||||||
5 蔟中および蛹の保護環境と各種の障害
A 温湿度による障害
日124号、日126号、日127号などに多発することのある半蛹(半化蛹)は稚蚕28℃、壮蚕22℃で軟葉を給与して飼育すると発生し易いが(Ⅱ2Aa、Ⅱ2Ba)、このような多発環境で飼育したものでも、蔟中の温湿度が低いと半蛹の発生が抑えられ、蔟、繭中の斃蚕も少ないと云う(第60表)。 なお、吐糸終了までを22℃に保護したものを28℃に移すと、全期間22℃保護のものに比べて半蛹の発生がやや多かったから、吐糸終了後の温度も幾分影響するものと考えられる(313)。
| 蚕品種 | 飼育条件 | 蔟中 | 化蛹蚕 歩合(%) |
半化蛹蚕 歩合(%) |
蔟・繭中斃蚕 歩合(%) |
||||
| 温度(℃) | 湿度(%) | 春 | 初秋 | 春 | 初秋 | 春 | 初秋 | ||
| 日124号 | 多発* | 28.0 28.0 25.0 25.3 22.5 22.3 |
80 82 85 80 56 61 |
10 25 72 |
0 14 77 |
35 55 13 |
48 43 8 |
54 18 14 |
41 42 14 |
| 少発** | 28.0 28.0 25.0 25.9 22.4 22.4 |
81 82 86 81 57 58 |
18 25 84 |
68 64 75 |
72 61 6 |
7 7 2 |
9 13 9 |
24 27 22 |
|
| 日126号 | 普通 | 28.0 27.8 28.0 27.8 21.8 22.0 22.4 22.0 |
80 83 55 56 89 90 58 57 |
28 40 55 75 |
53 75 81 84 |
54 50 31 12 |
15 5 7 3 |
18 9 13 13 |
31 19 12 12 |
| 日127号 | 普通 | 28.0 27.8 28.0 27.8 21.8 22.0 22.4 22.0 |
80 83 55 66 89 90 58 67 |
11 33 47 79 |
51 71 66 82 |
58 51 21 11 |
13 5 9 2 |
30 15 31 9 |
36 24 25 16 |
繭の縦作りもこの半蛹の発生を多くし、上蔟後60時間(24.8℃-25.5℃)縦にしておいたものは、その後横にしても矢張り半蛹が多かった(830)。 福田(108,110)の半蛹系統においても、湿度80%のとき、蔟中温度30℃では77.8%、25℃では27.8%、20℃では17.4%と、高温ほど半蛹の発生が多かったが。湿度は65-85%の間では半蛹の発生に影響をおよぼさなかった、95%では増大した。この系統では縦作り繭よりも横作り繭に半蛹が多く、前者47.1%、後者77.6%であった。この半蛹と佐藤らのものとの本質的な異同は明かでないが、福田のものは、脱皮殼を取除いてみると蛹皮が形成されており、また、その下には後に蛾皮が形成されるから、蛹化または成虫化ホルモンの異常によるものではなく、単に、脱皮過程が何らかの原因によって妨げられて生じるものと考えられている。仲野(660)の不吐糸蚕は蔟中温度が25.5℃のときよりも21.5℃のときに却って発生が多いと云う。
日127号、日126号、日124号などには、蔟中温湿度が高いと、ぼか繭、皿繭などの異常繭が発現し、日新にも僅かながらみられるが、日127号には特に多く、23℃-24℃、80%の蔟中環境においても発現することがある(565)。
上蔟以後の保護温度が高いと不受精卵が多くなるが、これがオキシキヌレニンの自働酸化によって着色して、死卵と見誤まられることがある(65,828)。
清水(842)によれば、蛹期間を通して30℃のような高温で保護すると、不受精卵や累積卵が多くなる。また普通には主として卵の裏面にだけ付着している膠着物質(Ⅰ2Ba)が卵の表面にも付着している。これは産卵の機構が異常になるためであろう。この膠着物質は、卵をエオシンの1%水溶液に浸すと赤く染まって容易に観察できる。発蛾前後に5℃に1週間以上冷蔵したものの卵にも同様なことが観察されるが、この場合には包卵被膜をかぶった未完成卵も産み出され、これは産卵後2、3日で潰れ卵になると云う。
三谷(539)は、高温(32℃、80%)と常温(23℃、80%)とに種繭を保護し、その蛾を互に掛け合わせて造卵数、産卵数および不受精卵数を調べ、また雄蛾の貯精嚢および交尾後の雌蛾の交尾嚢、受精嚢の肥大状況、精子の存否、ならびにこれらから取出した精子の運動力(大村法(751))などを調べて、高温保護が雄に悪影響をおよぼすことを確認した。
須貝・木口(870)は、化蛹後の雄蛹を種々な期間33℃で保護して更にこの点を追究し、蛹の後期に比べて前期が高温の影響を受け易いことを報告した。例えば、化蛹直後から23℃
で保護しておいた雄蛹を末期2日間だけ33℃においた場合には、正常雌蛾に交配すると約16%の不受精卵を出すに過ぎないが、末期6日間を33℃
におくと100%の不受精卵を出した(第18図)と云うのであるが、この図は不受精卵歩合の累積曲線であるから、時期別の感受性は明かでないように思われる。精子の量や運動力に異常があり、受精嚢にまで到達し得る精子のないこと、精室から脱出不能の精子が多いこと、脱出しても形態的に異常なものの多いことも観察された。なお無核精子(Ⅰ3Ab)の形成は高温によって著しく阻害されたが、これは無核精子の形成が主として蛹期に入ってから行なわれるためであろうと云う。
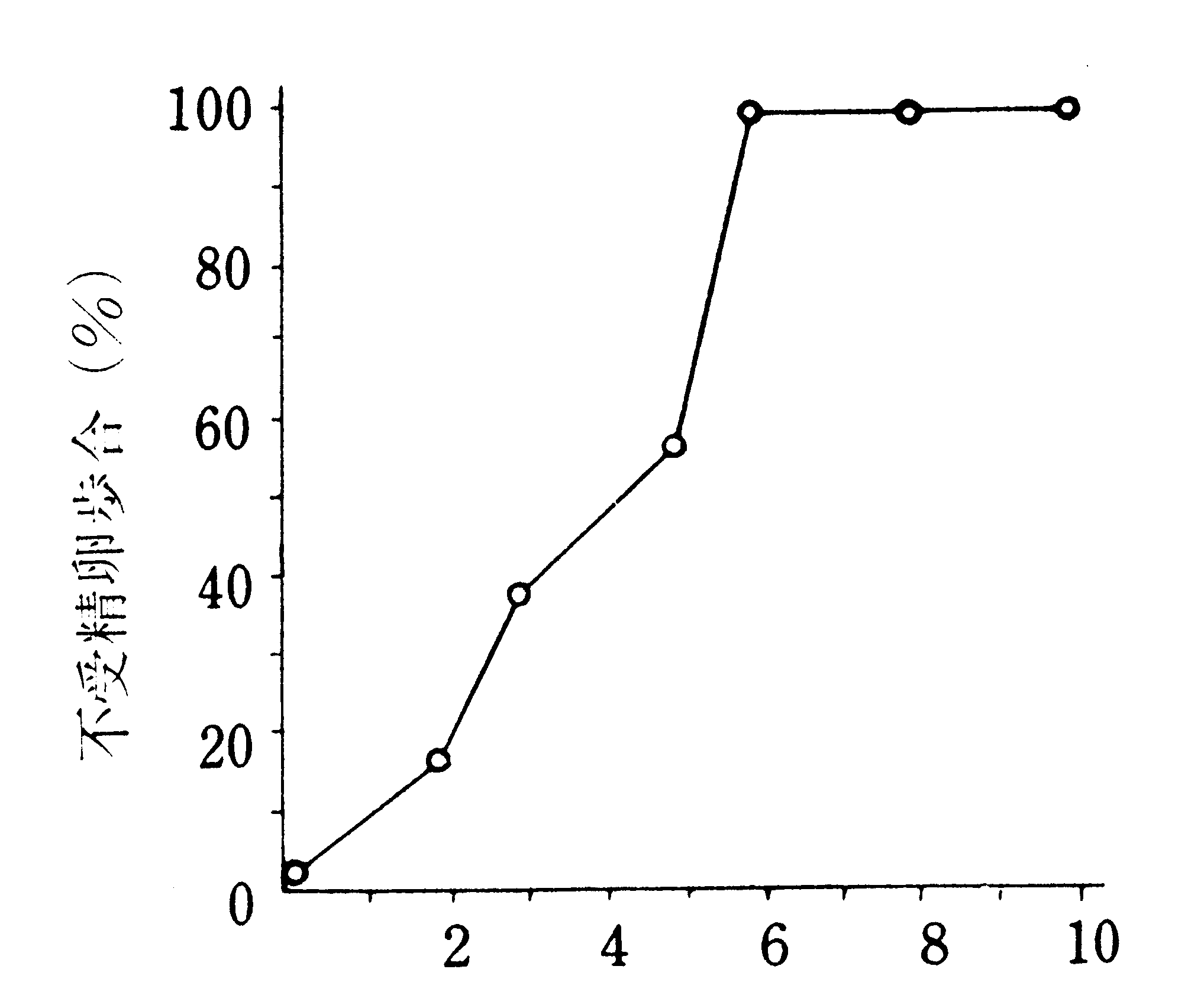
逆算した高温保護日数(日)
第18図 化蛾日から逆算した蛹の高温保護日数と不受精歩合(金色、33℃)(須貝・木口(870))
しかし、その後の研究によると(871)、化蛹後よりも、吐糸から化蛹までの前蛹期の方が高温に対して一層敏感で、雄の生殖減退を起こし易く、正常雌に交配したときの不受精卵歩合は、吐糸後1日間の32℃接触によって12.84%(23℃ の対照区では7.24%)、2日間の接触で29.00%、3日間で72.22%、4日間で89.52%にも達したが、32℃と23℃の接触を12時間々隔で交替すると、8日間続けても不受精卵歩合は対照区と変りがなかった。即ち、高温が244時間以上続くと雄の生殖減退が著しく高まるのである。有核精子の形成が行なわれている幼虫期の高温接触の影響をみるために、4令初期4日間、5令末期3日間の32℃接触も試験したが、雄による不受精卵歩合に影響がなかった。これは、給桑が十分であれば幼虫期の高温による雄の生殖減退が防止できることを示すものであろうと云う。蛹に45℃で8-24時間の高温度衝撃を与えた場合にも精子の運動カが阻害される(701)。
梅谷の陰茎付属筋肉の退化に基ずく不受精蛾の系統(1084,1122)においては、蛹の保護温度が高い(30℃)と異常蛾の発現が多く、しかも上蔟後30℃に保護したもののは5日目頃、20℃に保護したものは6日目頃の1日間だけがこの高温に対する感温期に当ると云う。
種繭を堆積すると内部の温度が高まる。例えば、外温は約18℃から漸次下って10時間後に約15℃になったにも拘らず、3貫500匁の生繭を布袋に詰め、大きさ70×40×30cmの竹龍に入れたものの中央部の温度は、この間に約5℃上昇し、外温との開きが約10℃に達したと云う測定結果もある(844)。外温の高いときはなお更であって、高温と共に炭酸ガス濃度をも高めて障害の原因になるから、繭の輸送および入荷した繭の処置には注意が必要である。
低温による障害については前節において述べたが、滝沢・勝野(1005a)、によれば、蛹期間を13℃で保護した蛾の睾丸は25℃で保護したものの睾丸よりも大きい。これは発育がよいためではなく、13℃保護のものの睾丸には説出し得ない未成熟精子が多く含まれているためであろうと云う。
B 炭酸ガスによる障害
炭酸ガス(濃度98%以上)中に一定時間密閉して死亡率を調べると、化蛹後の蚕は食桑中に比べて抵抗力が強い(第61表)。上蔟後、化蛹までのものは化蛹後のものよりも更に強く、この表と同様な試験の結果では死亡率が0であった。
| 蛹令 | 処理時間 | |||||||||||
| 1時間 | 3時間 | 5時間 | 10時間 | |||||||||
| 日1号 | 支4号 | 欧7号 | 日1号 | 支4号 | 欧7号 | 日1号 | 支4号 | 欧7号 | 日1号 | 支4号 | 欧7号 | |
化蛹1日目 2日目 3日目 4日目 5日目 6日目 7日目 8日目 9日目 10日目 11日目 12日目 |
% 0 10 10 0 0 0 10 0 0 10 10 0 |
% 60 40 20 10 0 0 0 0 0 0 10 - |
% 0 10 0 0 0 0 0 10 0 0 10 0 |
% 20 10 0 10 0 10 0 0 0 10 0 10 |
% 10 40 0 10 40 20 0 0 0 0 0 - |
% 0 0 0 10 0 30 0 0 0 0 0 0 |
% 20 10 0 20 0 0 10 10 0 0 0 10 |
% 30 10 40 10 0 0 0 0 0 10 0 - |
% 0 0 0 0 10 10 10 0 0 0 0 0 |
% 100 20 10 0 0 20 0 20 50 10 0 0 |
% 100 10 40 10 40 20 10 10 10 80 70 - |
% 40 10 0 20 50 20 40 40 20 20 50 0 |
しかし、このような蛹そのものの抵抗性とは別に、種繭の保護中に炭酸ガスの影響を受けたものの産卵には不受精卵が多く、その影響は蛹令の進むに連れて増大する(632)。
諸星(598)は、煉炭で補温して種繭を保護すると、不受精卵が数倍から十数倍にも増加する場合があり、その原因は炭酸ガスの影響と考えられると述べている。
煉炭の場合には、炭酸ガスのほかに亜硫酸ガスの影響も考えられる。また、農家が練炭を用いて不完全な補温をする場合には、管理に注意しないと1日の中の最高、最低の温度差が大きく、高温、低温、あるいは変温の害を受けるおそれがあり、これらは何れも産卵数の減少、不受精卵の増加の原因になる。