
第23図 蚕卵直後の卵の縦断模式
A:前極、D:背側、P:後極、V:腹側。
a:前極原形質、c:卵殻、d:連絡管
m:卵門丘、n:卵核、p:周辺細胞質
v:卵黄膜、y:卵黄
VI 蚕卵の形態、発生および生理
1 受精前の蚕卵の形態
家蚕の卵は、普通、偏平な小楕円体で、大きさは大約、長さ1.3mm、巾1.0mm、厚さ0.6mm(第42表)であるが、品種(697,982)、栄養(Ⅱ2Ac)、蛹の保護温度(Ⅲ3Bb)その他種々な条件によって変異をあらわす。また遺伝的な畸形も知られている(75,1037)。
産卵当時は乳白乃至黄色である。これは主として卵殻および卵黄膜を通してみえる卵黄の色によるものであるが、卵殻が着色しているものではその色も加わっている。
卵殻は最外層の卵膜で、均一な構造ではなく、多くの層を識別している報告もあるが、普通は外層、中層、内層および最内層に分けている(Ⅰ2Abii)。
卵殻の厚さは品種によって相違するが(568,697,916)、卵形を大きくする条件の下では卵殻も厚くなる。また卵の部分によってもかなり厚さが異なる(697)。最も厚いのは前極部(第23図)に徴小な臍状に突出している卵門丘の部分である。最も薄いのは卵門丘を取り巻く菊花状斑紋の周囲で、孵化の際に最初に喰破る個所はこの最も薄い部分に属している。
第23図 蚕卵直後の卵の縦断模式
A:前極、D:背側、P:後極、V:腹側。
a:前極原形質、c:卵殻、d:連絡管
m:卵門丘、n:卵核、p:周辺細胞質
v:卵黄膜、y:卵黄
外層の表面には無数の刻紋(卵紋)があるが、これは卵殻を分泌した包卵細胞の形のプリントされたものと考えられている(27,28,86,568,756)。刻紋の形は品種によって異なるから、蚕種の混同した場合などには判別の手がかりになる。卵門丘には外層を欠いている(29,960)。最内層は他の層と異なり、海綿状に近い構造のもののようである。卵殻の成分はキチンとかケラチンとか云う説もあったが、コリオニンである(567,719)。アミノ酸組成も調べられている(1057)。
卵門と気孔とが卵殻を内外に貫通している。卵門は前極部の卵門丘中央に開口している。1卵に1個であるが、開ロ部の直下で2-6本、普通3本の導管(精孔副枝)に分れて卵殻層内を通り(28,190,326)、その内端は長さ16-18μのラッパ状連絡管によって卵黄膜に連なっている。この連絡管は導管の延長であると云われているが(29)、導管末端部の形や、卵黄膜との接合部よりも導管との接合点で離れ易いことなどからみて、更に検討を要するように思われる。但し、卵黄膜のような膜状のものでないことは明かである。
卵門導管は、卵母細胞が7個の栄養細胞と連なっていた(Ⅰ2Aa)痕跡であるから、その数も7本であると云う説*があるが、少なくとも、 日本で普通にみる品種の卵には当てはまらない。桂(桂応祥 1934:広東特異蚕卵之形態与遺伝学的研究.仲愷農工学校研究報告 第2巻、第1号、1-260)の記載した広東産の蚕においても導管の数は3-4本が最も多く、1本、7本、8本のものは極く少ないと云う。
*朱洗・王高順・何家需・□詩宣 1954:家蚕混精雑交的研究(杉本行夫・亀井健三訳 1963:動物の多交配と遺伝性.新科学文献刊行会)
気孔は卵門丘を囲む菊花状斑紋部以外の卵殻全面に分布しているが(568)、刻紋との関係からみると、卵殻形成時に3個または4個の包卵細胞の接触点に当っていたと考えられる部分にのみ開口している(28,568,756)。従って、気孔の数は卵殻の分泌に関与した包卵細胞数のほぼ2倍に当る筈と考えられるが、大村・片岡(756)が5品種の卵について調べた気孔の数は1卵につき4661-9984個で、ほぼこの期待に一致すると云う。気孔の外面開ロ部の長径は3-4μであるが(30)、内方ほど細く、縦断面は漏斗状を呈する。
蚕卵を色素の水溶液に浸すと卵殻の中層および内層まで染色されるから(916,1070)、水溶液が卵殻層内に侵入することは確かであるが、最内層を通過して卵内にまで入るか否かは明かでなかった。 しかし、pH4.2の亜硝酸カリ(KN02)水溶液に蚕卵を浸すと極めて死亡率が高いので、これが郭内に容易に侵入することは確かである(709,1051)。ただ、亜硝酸カリの溶液に蔗糖を加えて浸透圧を高めると卵の生存率が高まったと云われているのは、2回の試験のうち1回だけがそのような結果で、他の1回は反対の結果を示していることからみてなお検討を要する。卵殻を貫通している気孔その他の細孔には空気が入っているので、浸透圧は働きにくい条件になっていると思われるが、実際に、濃厚な塩水に浸しても内容の収縮は起こらない。またN02が卵殻を通過したか否かをα-ナフチルアミンとスルファニル酸との混合液で定性してみると、亜硝酸イオンの卵殻通過は蔗糖の添加によっては影響を受けず、pHの影響の大きいことがわかった(未発表)。 pH5.6の溶液では蚕卵に対する亜硝酸カリの害がないと云われているが、この溶液の場合には亜硝酸イオンの卵殻通過をこの定性で確認することができなかった。なお蚕卵は、数カ月間水に浸しておいても、水を吸って膨潤し破裂すると云うようなことはない(Ⅵ3Bb)。
卵黄膜は卵母細胞の周辺細胞質から形成される無構造な膜である(496,501)。卵を生のまま生理食塩水中で解剖して漿膜に包まれた内容を取出すと、卵黄膜は卵殻内面に付着して残るが、卵を固定してから卵殻を除去すると卵黄膜は漿膜面に付着してとれてくる(未発表)。孵化の際には卵殻の内面に付着して残る(387a,983)。
卵黄膜の内部には原形質および卵黄が充満している。卵黄には脂肪性顆粒と蛋自性顆粒とが区別せられ(Ⅰ2Abii)、固定切片でみると、直径が前者は3.0μ(1.4-4.6μ)、後者は6.0μ(1.5-9.8μ)ぐらいである(290,496,501)。
卵黄膜の直下に卵黄顆粒を含まない細胞質の薄層(周辺細胞質)がある(555)。これは、受精後に分裂核が入って増殖、分化する部分で、胚発生上重要な意味を持っている。
卵門の付近で周辺細胞質の層の特別に厚くなった部分があり、前極細胞質と呼ばれることもあるが、産卵当時の卵核はこの部分に含まれている。交尾して順調に産み出された卵においては、産卵当時の卵核は卵母細胞の第一成熟分裂の後期の初めの状態であるが、未交尾蛾の産卵においては第一成熟分裂の中期であると云う(813-815)。
圧し潰しに対する蚕卵の硬さは主として卵殻の厚さによってきまる。硬さは卵の大小によってきまると云う報告もあるが、これは同一品種では、小さい卵が一般に卵殻も薄いためである。しかし、圧し潰しに対する硬さは、卵殻の一点の強さだけによるものではなく、卵全体としての構造が関係するから、小さい卵が必ずしも潰れ易いとは限らない(1,362,876,1079,1080)。高須(967)は、蚕卵の硬さは飼育時期、用桑その他の卵殻の厚さに影響すると考えられる諸条件によって相違するほか、催青の進むに連れて硬度が減じ、点青乃至催青期において最も潰れ易くなるが、孵化直前にはかなり回復することなどからみて、卵殻ばかりではなく卵内容物質の変化とも密接な関係があるものと考えている。
2 発生
A 受精
卵門から精子の侵入するのは卵が母体をはなれる数秒前である(Ⅰ2Ba)。精子の侵入によって第一に変化するのは周辺細胞質である。堅い卵殻で覆われている蚕卵においては、ウニなどにおけるようなこまかい観察や実験を行なうことはできないが、少なくとも周辺細胞質の表層に変化が起こって(ゲル化であろう)、表層と卵黄膜とが離れ易くなることは確かである(387a)。末交尾蛾の卵に比べて、交尾蛾の卵においては産卵当時の成熟分裂の進んでいるのも、精子侵入の刺戟によるものと考えられる。
精子の尾部は非常に長く、卵の長さの半分ぐらいもあるが(Ⅰ3Ab)、これが全部卵内に入ってしまうのか、精子の頭部が卵の周辺細胞質に達した処で尾部から離れるのかは確認されていない。盛んに産卵している交尾蛾を生理食塩水中で手早く解剖し、丁度、膣前庭にある卵を取出してみると、精子の長い尾部が卵門から出ているのを観察することができるが、前庭より下った位置の卵ではこれをみることができない。尾部が侵入してしまうのか、粘液腺からの膠着物が付与されるためにこれと一緒になってしまうのかは不明である。
1個の卵に侵入する精子の数は1個より多いのが普通で、11個入っていた例も報告されている(325,759,814,1074)。朱洗ら(前節参照)は、卵門導管が7本であるから、侵入する粒子数は最高7個であると云っているが、導管数によって制限は受けないもののようである。
1卵内に多数の精子が入っても、卵核と結合し得るのは普通の場合は1個の精子だけであるが、母親遺伝をするモザイク系統の雌蛾に、目標になる優性遺伝子ホモの雄蛾と劣性遺伝子ホモの雄蛾とを二重交尾させてみると、1個だけではなく他の精子も受精力を保持していることがわかる。母親遺伝をするモザイク系統の卵においては、正常の雌性前核のほかに1個の極体も別の精核と結合して受精する。しかし、こうしてできた分裂核は統合して1個体を作るので、二つの受精に由来する違った性質が体の部分に別々に発現して、モザイクや雌雄モザイク蚕になるのである(1046)。
このような試験結果から興味のあるのは、二重交尾をさせた2頭の雄蛾の精子は、交尾の順序に従って使用されるのではなく、混合して1卵内に侵入し得ることである。これは普通の系統の蚕においても同様で、1頭の雌蛾に優性遺伝子ホモの雄蛾と劣性遺伝子ホモの雄蛾とを二重または三重に交尾させてみても、初めに交尾させた雄蛾の子が特別に多いとは限らないから、受精嚢に入った精子は交尾の前後に拘らず混合使用されるものと考えられる。従って、このような交雑の場合に、交尾の順序を変えても優性の子供と劣性の子供との割合が同じ方向に片寄るようであれば、両雄蛾の精子の間に受精競争が行なわれ、受精力の強い精子が多くの卵を受精させたものと考えなければならない(1002,1003,1007,1008)。大村(753)によれば、幼虫期に出血させたり、蛹または蛾で長く冷蔵した雄蛾の精子は受精競走において無処理のものに劣ると云う。受精競走はその結果からみて選択受精とも云う。小村(708)は二重交尾をさせた場合における、両雄蛾それぞれの性質を受けた卵の産卵順序を調べている。
卵内に入った精子の核は卵核と結合するばかりでなく、条件によっては精核同志で結合して胚またはその部分を作ることがある。橋本(176)は、交尾蛾の産んだ産卵後1時間以内の卵に36.5-41.5℃の高温を1時間作用させると、精核同志の結合によって生じたものと考えなければ遺伝学的に説明し得ない個体や、モザイク蚕の現われることを報告したが、田島はこの現象を2精子メロゴニーと呼び、多くの実験を行なった(1046)。但し、メロゴニーと云う言葉がついていると本来のメロゴニー(卵片発生)とまぎらわしいので、最近では雄核発生(androgenesis)と呼ぶことが多い。
高温処理による精核同志の結合は細胞学的にも確められている(816)。交尾蛾の産んだ卵を高温で処理すると単為発生よりも雄核発生の起こり易いのは、精核に比べて卵核が高温や低温に対して弱いことが原因と考えられるが(181-183,819)、産卵後3時間目頃に処理卵の前極部付近をみると、卵の長径の1/5ぐらいの楕円圏内において精核が活発に運動しているから、これも精核の結合を前けるのであろうと云う(1030)。 雄核発生は低温処理(1013,1014)、炭酸ガス処理(1050,1052,1053)、などによっても起こる。
未交尾蛾の産卵に種々な刺戟を与えて単為発生を行なわせる実験(327-329,813,815-817,821)、蛾または化蛾に近付いた蛹の卵巣内から取出した卵の温湯処理による単為発生(70,179,180,204,210,211,896、897)などに関しても多くの報告がある。卵巣から取出した卵を処理して得られる単為発生蚕は全部雄である。これは第一成熟分裂において染色体が両極に分かれないためと考えられる(70,817)。交尾蛾の卵は産卵直後に処理しても、卵核だけが単独で発生することは極めて少ないと云う(181)。
未交尾蛾の、特別な刺戟を与えない自然のままの産卵においても、胚が単為発生によって発育を始めることは珍らしくない。不受精卵と云われているものを切片にして調べると、漿膜の不完全に形成されているもの、異常な胚盤形成の認められるもの、胚ができて死んでいるものなど、種々な発育階程の卵の含まれていることが観察される(828,1074)。これは人為単為発生の成功しなかった卵についても同様である(812)。ただ、自然単為発生においては孵化にまで達するものが極めて少ない(327)。
佐藤(813,814)によれば、精子の侵入した卵においては、80゚F(26.6℃)の場合には産卵後5分内外で第一成熟分裂が完了し、第二成熟分裂は産卵後60分頃から始まって80分頃に終り、雌雄両前核は産卵後2時間目頃に合体(受精)して第一分裂核を作る。第一分裂核はその後1時間以内に最初の分裂を起こすと云う。
これに対し、田島ら(1052,1053)は、適当な卵色遺伝子を持たせた雌雄を交配し、産卵後、0-20分の卵から始め、以後20分毎に一定時間の炭酸ガス処理を施こし、雄核発生その他の例外型および死卵の発現を調査した結果から、第一成熟分裂、第二成熟分裂、分裂核の第1回および第2回目の分裂完了時を、それぞれ、産卵後50、110、190および250分頃と推定した。村上・鬼丸(601)が蚕卵を圧し潰して細胞学的に調べた結果では、分裂核の分裂週期は40-50分と考えられたが、その後、X線による蚕卵致死率の変動から村上(600,602)が推定した分裂核の分裂核週期は60分である。大槻・村上(774a)の最近の報告によれば、第一成熟分裂は産卵後40分頃に終了し、分裂核の分裂は、少なくとも第3回までは同調で、周期は60分であった。
朱ら(Ⅵ1)は、二化性種の越年性卵においては、極体が卵膜(受精模であると云うが詳細不明)の外に放出されることが多く、極体を膜外に放出した卵は不越年性になるから、この放出が越年、不越年の目印になると云い、浸酸処理はこの極体を人為的に放出させて、不越年にする方法であると述べているが、極体の形成されるのは上記のように産卵後間もない時期であり、浸酸処理を行なうのは普通にはこれよりも約20時間後であるから、極体の追跡は困難になっている筈である(814)。
B 胚の発有階程
胚発生の知識が産業に直接結び付いていると云う点では蚕は他に比類のない動物と云えるが、蚕糸と云う立場だけから考えられたためか、最も得易い材料を極めて多数の人が手がけている割には、正しい意味での昆虫発生学の材料として用いられることの少なかったのも蚕である。
高見・北沢(935)は、蚕種保護の実用のために、蚕の胚発育を次ぎの30段階に分けている。
┌Stage 1 受精期
│Stage 2 分割期
│Stage 3 胚分化期
前休眠期 ─┤Stage 4 卵黄分割期
│Stage 5 だるま形期
│Stage 6 こけし形期
└Stage 7 へら形期
┌Stage 8 休眠Ⅰ期
休眠期 ─┴Stage 9 休眠Ⅱ期
┌Stage 10 前甲期
│Stage 11 甲期
越冬期 ─┤Stage 12 乙A期
└Stage 13 乙B期
┌Stage 14 丙A期
臨界期 ─┴Stage 15 丙B期
┌Stage 16 丁A期
│Stage 17 丁B期
│Stage 18 上唇突起発現期
│Stage 19 短縮期
│Stage 20 頭胸分節期
器官形成期─┤Stage 21 反転期
│Stage 22 反転完了期
│Stage 23 毛瘤発生期
│Stage 24 剛毛発生期
└Stage 25 気管螺旋糸発生期
┌Stage 26 点青Ⅰ期
│Stage 27 点青Ⅱ期
完成期* ─┤Stage 28 催青Ⅰ期
│Stage 29 催青Ⅱ期
└Stage 30 孵化期
*原著においては気管形成期に入れていた。
このうち、蚕種保護のために特に必要な階程だけを、特徴を強調して描いたのが第24図である。
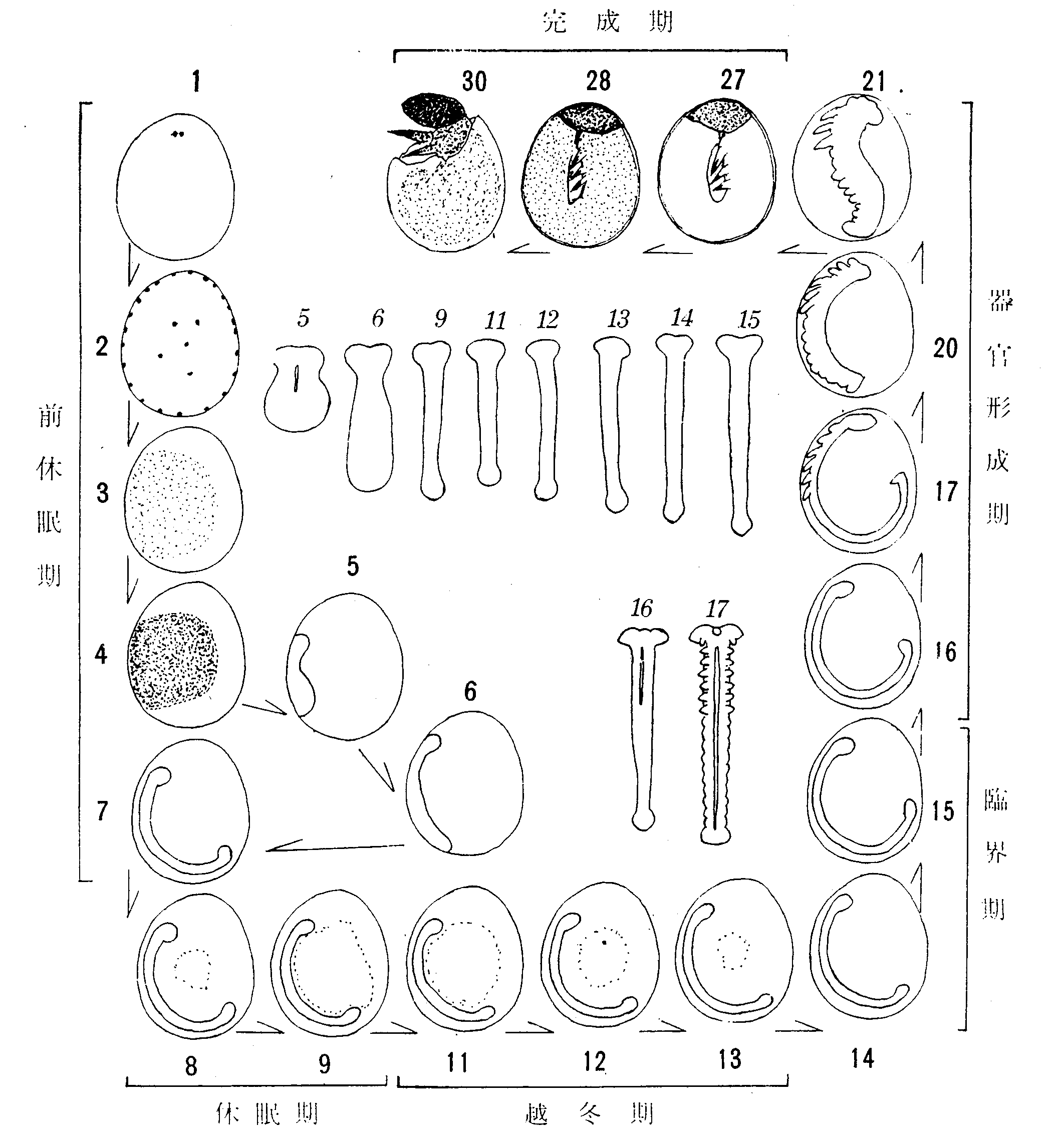
第24図 越年性卵の胚発生模式
ゴチックの数字はその発生段階(本文参照)の卵の縦断または透視。
イタリックの数字はその段階の胚を取出して腹面からみた輪郭である。
以下これによって解説する。発生経過は23-25℃における産卵後の時間である。
a 前休眠期
Stage 1 受精期 第二成熟分裂を終えた卵核(雌性前核)は卵内のほぼ黒点で示した位置で精核(雄性前核)と合体する。2時間目頃である。
Stage 2 分割期 受精によって生じた第一分裂核は3時間目頃から分裂を始め、以後、娘核は分裂を繰返えしながら周辺部に向って移動し、10時間目頃に周辺細胞質中に入り込み、12時間目頃にはこに1層の細胞層(胚盤)を作る(289,291,560,561)。一部の分裂核は卵黄細胞となって卵内に残る。
分裂核の分裂週期を1時間とすると、同調分裂が続けば8-9回の分裂で胚盤期に入る訳で、EphestiaやDrosophilaなどで知られているのとほぼ同じであるが、蚕卵においては何時まで同調分裂が続くのか正確にはわかっていない。
Stage 3 胚分化期 15-20時間の時期である。 15時間目頃から腹側を中心にして胚盤の広い範囲が厚みを増し、この部分が胚に発育するから、後に胚になる部分(胚域)と漿膜や羊膜に発達する部分(胚外域)とが識別できるようになる。識別できるようになった胚域は胚帯と呼ばれている。
蚕の卵は、一見、偏平な楕円体であるが、相称形ではなく、第23図に示すように尖りめの部分(前極)、その反対側の後極、膨らみのある腹側、その反対側の背側が識別できるが、胚は前極に頭部を向け、腹側に腹面を向けて作られるのであって(911)、正常な発生においてはこの向きが変わることは決してない。そればかりではなく、胚域のどの部分が、将来、胚のどの部分になるかと云うことも、胚域がはっきりしてくる以前の胚盤期、または分裂核が周辺細胞質に到着するより前から、既にかなりの程度に決定している(918,919,922)。宮が卵の種々な部分を焼き殺して生殖腺の発生を調べたのは(Ⅰ1)、この前決定の性質(484)を利用した研究である。
雌雄によって、催青卵期の卵の中での胚の右向き、左向きが違うと云う説があるが(ⅣEc)、卵の中で胚の形成されるのが腹側にきまっていること、産み出される途中の卵の廻転によってこの腹側が右になるか左になるかが偶然にきまり、反転によって胚の左右位が形成のときとは反対になることなどを考えれば、この説には理論上からも根拠のないことがわかる。
Stage 4 卵黄分割期 胚帯は腹側に向って、左右から(図で云えば表裏から)収縮してますます明瞭となり、20時間目頃には腹側を中心にして馬に鞍をかけたような形になる。
卵黄は、この頃までは蛋自性顆粒と脂肪性顆粒との単なる集積に過ぎないが、やがて卵黄細胞を含んだ小塊に切崩され初める(289,291,947,1157)。これを卵黄分割と云うが、卵黄分割は卵の周辺部から始まって次第に中央部におよび、胚の直下にも進行する。分割の結果、卵黄は卵黄細胞の含有物の観を呈するようになるが、この分割体(卵黄細胞)は卵黄粒団と呼ばれることもある。卵黄分割が完了すると、胚に卵黄が粘着しなくなるため、生で解剖した場合に胚が離れ易くなる。越年性卵の卵黄細胞においては脂肪性顆粒が核の周囲に集積しており、不越年性卵においてはこれが細胞内蛋白顆粒の間に分散している(926)。
この間に胚外域の細胞が胚帯表面を覆うように延びて漿膜と羊膜とを作る(238,904,921,1073)。漿膜細胞の大きさおよび数は品種によって異なり、母親遺伝をすると云う(333)。足立(15)は、産卵後40時間になっても、なお卵内から分裂核が卵の表層に到着して、既に着色した漿膜の形成に参加するように述べているが、そのようなことはない。
加温即時浸酸の適期はこの時期(20時間目)である。
この頃までの胚の発育は越年性卵でも不越年卵でも大差はないが、その後急に違いが現われてくる。外見に現われる越年性卵の最も著しい変化は、1日余りで漿膜細胞に色素形成が始まり、卵が淡紅褐色に色付いてくることである(332,1132)。卵の酸素消費量もこの頃から急に低下し(Ⅵ3B)、グリコーゲンの含量が減少する(Ⅴ2B)。
Stage 5 だるま形期 胚帯収縮が更に進むと左右から縊れが入り、胚の形に頭部と尾部との区別ができてくる。“だるま”形とはその形から名付けられたのであって、越年性卵においては約30時間、不越年性卵においては約25時間の頃である。
Stage 6 こけし形期 越年性卵の35時間目頃に当る。20時間目(Stage 4)に加温即時浸酸を施した卵は、条件によっても異なるが、浸酸後約18時間でこの時期になる。
即時浸酸種の冷蔵は18時間後に行なうのがよいとし、俗にダルマ期冷蔵と云われているが、即浸後18時間の胚は、だるま形よりも長くなった“こけし”形の時期であって(913,917,933)、真のだるま形期(Stage 5)に冷蔵すると成績が悪い(Ⅷ2Cb)。
完成した漿膜の表面には、普通気付かれないでいるが、薄い漿膜クチクラ(serosal cuticle)がある。これは36-48時間目頃に漿膜細胞から分泌される(556)。漿膜クチクラの形成と空気や液体の透過性との関係はわかっていない。
Stage 7 ヘら期形 越年性卵の3日目である。以後、休眠中を通じて、胚の外形は殆ど変らない。3日までは、胚を構成する細胞に分裂中のものがあるが、4日になると分裂はみられなくなる(386)。従って、越年性卵の胚は産卵後約3日で前休眠期の成長を完了するものと考えられる。
卵色は日に日に濃くなり、1週間ぐらいでその品種固有の色に落付く。卵内の3・ヒドロキシキヌレニンはこの色素形成のために消費されて、産卵後1日ぐらいから急激に減少するが、5-8日で減少が殆ど停止する(244,1132)。越年性の完成が不良な場合にはこの消費が少なく、また即時浸酸種においては越年性卵に比べて消費が少ない(355,357)。卵色は漿漠の色と卵殻の色、ときには透けてみえる卵黄の色もこれに加わった複合色である。越年性卵でも漿膜の着色しない突然変異型がある。
b 休眠期
水野の休眠期(Ⅵ2Bc)とは違う。何時からが休眠期であるのか、はっきりした区切りはできないが、2週間ぐらいで、一応、休眠と云われる状態になる(Ⅴ1Ba)。休眠Ⅰと休眠Ⅱも明確な区切りをつけることはできない。
Stage 8 休眠Ⅰ期 休眠期の胚の形は品種によってかなり相違する。前休眠期との相違で著しいのは胚の形よりも卵黄の状態の変化である。生体で解剖してみると、前休眠期には卵黄細胞が互に遊離しており、胚に付着しているものも離れ易いが、次第に離れにくくなる。同時に、中央部にある卵黄細胞が周辺部に向って移動勤し、集塊となって胚および漿膜に固着するため、卵の中央部に卵黄のない部分ができてくる(289,291,928,946,1078,1157)。この部分は次第に大きくなり、透過光線で透けて明るくみえるようになる。
Stage 9 休眠Ⅱ期 卵の中央の卵黄を欠く部分はますます拡大し、卵黄に堅く包まれた胚は、生体解剖では卵黄との見分けが困難な状態になる。
越年性卵はこのような状態で発育休止のまま夏秋期を経過する。卵黄の色は体液の色によって異なり、黄血蚕の卵は産卵当時から濃い黄色であるが、これとは違い、色素を形成して次第に着色するものがある。第2白卵(w2)種の卵が産卵後次第に橙色に着色するのは、漿膜にも若干色素ができるが、主として卵黄の着色によるものである。カンボージュなどの多化蚕の卵の着色にも同様のことがみられる。赤卵(re)の卵黄も橙色に着色する(652)。詳細に調べると、正常卵色の卵においても、卵内に点々と紅色の色素粒を形成しているものが珍らしくない(385)。
c 越冬期
自然湿度で保護してある蚕種の胚には、東京地方では12月初句になると、ぼつぼつと細胞分裂が現われる。しかし、その数は少なく、これを25℃で催青しても1ヵ月では孵化のみられないことが珍らしくない(386)。蚕の越年性卵は完全な休眠状態ではなく、このように一部休眠の醒め初めた状態で冬を迎えるのであって、低温に接触しなければ揃って発育するようにならないので、三浦はこの時期を対寒的発育期と呼んでいる(546,552)。
Stage 10 前甲期 催青すると胚が発育するから、休眠は脱しているが、完全に活性化した甲期とは違い、孵化までに非常に長い日数を要し、また発育が不揃いである。漿膜に堅くはり付いていた胚や卵黄が幾分ゆるんではいるが、形態的に休眠期とはっきり区別のできる特徴はない。第24図には省略した。
Stage 11 甲期 水野(577,580)の甲胚子に当る。甲胚子を水野が“所謂休眠期の時である”と記載しているために、今日の言葉での休眠と混同して、甲胚子は休眠胚であると誤解されていることがある。しかし、水野の云う休眠期は“休晩期に入りたる胚子は其後温暖に逢うことによって発育を始むるものである”と述べていること、および“甲胚子を摂氏17度半(華氏63度半)に1日間接触すれば凡そ乙A胚子を得る”と記述していることからわかるように、未だ発育を始めてはいないが胚が十分に活性化して、完全な発育を開始し得る状態に達した時期を指すものである。今日では越年性卵の胚が発育休止状態になることを休眠に入ると云うが、水野はこの時期に対しては休眠と云う言葉を使っていない。言葉は同じでも、当時の休眠期(水野以外にも同様の記載がある)は今日の休眠解消期に当るから、注意しなければならない。
また、水野の発生段階においては、甲胚子には中胚葉塊の環節的配列が認められない、と解説されているが、家蚕の胚には、前休眠期の終りから休眠期、越冬期、臨界期を通じて、中胚葉の環節状の配列が認められない時期はない。ただ、甲の頃は胚が短縮し、中胚葉塊が互に密接していて認めにくいだけである。この時期の中胚葉塊の形は品種によって異なる。どの部分のものがきまって大きいと云うこともない。
この時期の胚の形として、頭部の巾が広く、尾部ほど細くなって、頭葉と顎部との間に縊れのない図の示されていることが多いが、品種や標本の作り方によっても異なり、常にこのような形とは限らない。結局、完全に活性化はしているが未だ長さに伸びの認められない時期が甲と云うことになる。
生体解剖で観察すると、一部の卵黄細胞が遊離しているが、未だ無定形な集塊状を呈するものが多い。
Stage 12 乙A期 水野の乙Aに当る。甲より幾分長くなり、中胚葉塊相互の間隔がやや開き、胸節のものが他の部分のものに比べて幾分大きいが、明確な区別点はない。遊離の卵黄細胞が増加するが、これも連続的な変化に過ぎない。
Stage 13 乙B期 水野の乙Bに当る。胸節の中胚葉塊が他の部分のものに比べてはっきり大きくなるが、未だ体巾一杯には拡がっていない。卵黄細胞は大部分遊離するが、外形のすっりきりしないものが多く、大塊を作っているものもある。乙Aとの違いは連続的である。頭葉が大きくなり、体長も長いが、量的な違いで、乙Aとの聞の明確な区別点にはならない。
越冬期の胚には、暖い日が続くと細胞分裂の認められることがあるが、5℃以下の温度に保護されているものにおいては細胞分裂はみられない。この時期の胚発育は、休眠による透過性の変化と冬期の低温とによって脱水状態になっていた胚および卵黄細胞が、活性化に伴なって水分を取りもどし、伸びだした状態と考えられる(386,945)。甲は透通性が回復して伸び得る状態になったとき、乙Aはこれが伸び初めたとき、乙Bは伸長の終りに近い時期とみるべきもの(Ⅵ3Da)であるから、基準を設けてこれらの間をはっきり区切ることはむずかしい。蚕種保護の上からも、これらの時期の胚を形態によって区別することに苦心するよりは、胚が完全に活性化して、温度を上げれば順調に発育し得る状態になったときを確認することが大切である。水野が、17.5℃に1日おけば甲が几そ乙Aになり、翌日乙B、翌々日に丙A、更に1日で丙Bになると記載していることは、この意味で非常に含みのあることと考見なければならない。
d 臨界期
東京付近の自然温度においては、2月末から3月初めにかけてみられる越年種の状態である。頭葉が左右に開き、越冬期には体の中心線に沿って塊状に近い形で並んでいた中胚葉が左右に拡がり、18個の体節がかぞえられる。卵黄細胞は完全に遊離して卵内一様に分布し、卵の中央部の、透過光線で透けてみえた部分がなくなる。
越冬期から引続き蚕種を5℃に保護しておくと、胚はその温度の下で臨界期までは発育し、100日ぐらいおけば細胞分裂も現われるが、2.5℃では活性化はするが250日たっても臨界期の形態を示すに至らなかった(945)。
三浦(546,552)は、寒冷に接触させる必要がなくなり、温度の上昇に連れて胚が発育する状態になったとして、この時期以後を対暖的発育期と呼んでいる。
Stage 14 丙A期 水野の丙Aに当る。頭葉が開いて大きく、体は伸びて細長な感じになる。特に体後半の伸長が著しい。中胚葉塊は左右に拡がって帯状を呈する。卵黄細胞は完全に遊離する。
Stage 15 丙B期 水野の丙Bに当る。頭葉は十分に開き、倒三角形を呈する。胸節の中胚葉塊は左右に分れる直前で、先端で付着し合ったハの字形を呈する場合が多い。尾節は後端が尖り菱形に近くなる。水野は胸部の1、2節に微かんに正中線の現われたときまでを丙Bとしているが、本書においては正中線(神経溝である)の現われたものはStage 16 とする。
細胞分裂を伴なわずに伸長し得る極限の階程で、これ以後の発育は細胞分裂を件わなければ進行しない(386)。休眠期および越冬期の胚を生体で生理食塩水中に取出すと、著しい屈曲、捻転をあらわすが、この時期になると屈曲は全く起こらない。胚の吸水伸長の完了したことを示すものであろう。
水野は丙Bを胚の発育経過中で最長の時期であるとして最長期胚子と名付けたが、実際にはこの時期よりも次ぎの丁A、丁B期の胚の方が長いから、梅谷(1101)は水野の丙Bを細長期、水野の丁Aを最長期と呼んでいる。岩崎は丙Bを臨界期と呼んでいる。呼び名が混乱しているから、古い文献を読む際には注意が必要であるが、普通には、最長期と云えば丙B期のことである。
蚕種の保護、取扱いは胚の発育階程を基礎にして行なうと云われているが、その発育階程のうちで最も重要なのはこの丙Bの時期でる。単式冷蔵においても中間手入れ方式の冷蔵においても、この時期まで胚を発育させて冷蔵するのが越年種の冬期保護の常法である。この時期を過ぎると冷蔵に耐える力が低下するから、鑑定を誤まらないように注意しなければならならない。上に述べたように、越冬期の胚の鑑定には形態的に明確な基準が乏しいが、越冬期の胚を調べるのは、丙Bの鑑定を誤まらないための補助的な調査であると云ってもよいぐらいで、蚕種保護上大切なのは丙Bである。
中胚葉が左右に拡がって、胸節においては、ハの宇の上端が付着し合ったような形になるが、それが未だ左右に分れない時期が丙Bで、体の正中線に神経溝がみえ初め(胸節から始まる)たものは既にこの時期を過ぎている。胚の形は品種によっても異なるから、大きさとか、長さとか、頭葉の形とかなどのように変異の多いものよりも、15℃で保護した場合に、神経溝のみえ初める1日前、付属肢の現われ初める2日前と云うことを基準にして、丙Bの形に慣れ
ることが必要である。
ただ注意しなければならないのは、丙B期の胚が2.5℃では100日、5℃では60日の冷蔵に耐えると云われているのは、活性化と胚の発育とが調和して進行している場合には、胚が丙Bになったときの生理状態が冷蔵に適していると云う意味であって、条件が違えばこの形態と生理状態との間にずれが生じ、丙Bが必らずしも冷蔵適期を示さないことに注意する必要がある(Ⅶ1Da)。丙B期が重要であると云うのは、丙Bと云う形態を通して推定される生理状態が重要なのであって、形態が重要なのではない。
e 器官形成期
胚に各種の器官の形成される時期である。
Stage 16 丁A期 胚の正中線に神経溝が現われ、頭葉が角ばってくる。水野の丁Aの時期である。上に述べたように、梅谷はこの時期を最長期と名付けたが、実際には顎節および胸節に付属肢の生じた頃(丁B)の胚の方が長い。もっとも、梅谷のF胚子(水野の丁Aに相当すると云う)には“上顎、下顋、下唇の3対の突起および胸脚の3対の突起徴かに認め得”られると云うから(1101)、水野の丁Aよりは進んだ時期であって、ここにもまた混乱がある。
Stage 17 丁B期 顎節および胸節に付属肢が発生する。続いて腹節にも突起が現われる。水野の丁Bに当る。胚は反転期以前では最も長いが、胚発育期間を通じて云えば、最も長いのは孵化直前の完成胚であるから、最長期と云う名称はこの時期に対しても適当ではない。
不越年性卵の胚は産卵後2日でこの階程になる。
Stage 18 上唇突起発現期 上唇突起は丁A期から発生が始まるが、これが次第に前方に向って移動し、2個の突起となって頭端に現われるのがこの時期である。この期の初めには尾節の巾が腹部後端の巾より広いが、次第に狭ばまり、後期には両者の巾が等しくなる。ほぼ水野の戊Aに相当すると考えられる。第24図には省略した。不越年性卵は産卵後2日半でこの時期になる。
Stage 19 短縮期 体長目立って短縮、尾節が腹部の後端より狭く、小さくなる。気門および絹糸腺陥入。ほぼ水野の戊Bに相当すると考えられる。不越年卵は3日でこの時期になる。第24図には省略。
Stage 20 頭胸分節期 体長は著しく短縮する。顎節が次第に集合し、将来の頭部になる部分と胸部になる部分ととの区別がつくようになる。触肢突起が前進して口陥と同列になる。尾節は非常に小さい。ほぼ水野の戊Cに当る。不越年性卵の3日半。
Stage 21 反転期 これまで卵周に沿って卵の腹側に位置を占め、背面に湾曲していた胚に湾曲転換が起こり、同時に胚が背側へ転位する運動の全期間で、ほぼ水野の己Aに当る。不越年性卵の4-5日目である。ヘロルド腺発生(668)。
中田(668)は、第1腹節から後方の体節が卵周を離れて内部に移動し始めた時期を反転前期第1期、第1腹節から後方が殆ど一直線になった時期を反転前期第2期、第7腹節から後方が腹面湾曲に転じた時期を反転後期第1期(第24図、第25図a)、第1腹節から後方が脱脂湾曲に転じた時期を反転後期第2期(第25図b)に細別している。
反転の機構については諸説がある(60,145,268,484,920,1100)。
反転すると直ぐ胚脱皮が起こる(703)。脱皮は引き続いて2回行なわれる(未発表)。前胸腺の働きとは関係がない(951)。
Stage 22 反転完了期 湾曲転換が完了し、口部の諸器官はロ陥の周囲に集まり、ほぼ蟻蚕にみられるような配置をとる(第25図c)。不越年性卵の5日目。
Stage 23 毛瘤発生期 毛瘤発生。尾角が生じる。ほぼ水野の己Bに該当すると思われる(第25図d)。不越年性卵の6日目。この頃から胚に脈博が認められる(1197)。
Stege 24 剛毛発生期 剛毛発生。胸肢に爪が生じ、大顎に鋸歯形成。単眼の着色始まる。図省略。不越年性卵の6日半。
Stage 25 気管螺旋糸発生期 気管に螺旋糸発生、大顎着色(第25図e)。不越年性卵の7日目。
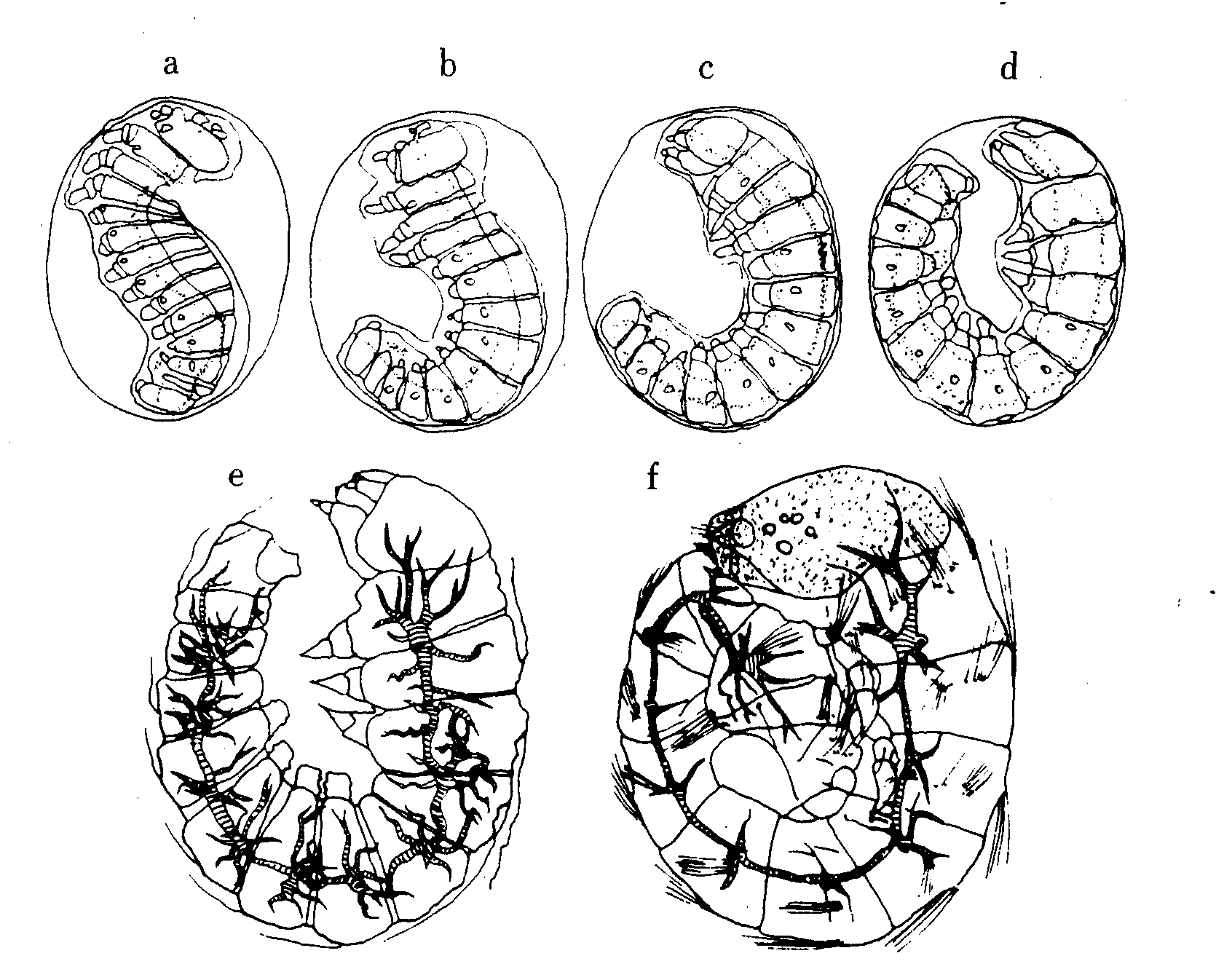
第25図 反転後期から点青初期までの胚(中田)(668)
a:Stage21 反転後期第1期、b:Stage21 反転後期第2期
c:Stage22 反転完了期、d:Stage23 毛瘤発生期
e:Stage25 気管螺旋糸発生期、f:Stage26 点青Ⅰ期
f 完成期
胚体の完成する時期である。生体で解剖してみると、取出した胚は生理食塩水中で活発に体を動かす。
Stage 26 点青Ⅰ期 胚は未だ漿膜を飲み込んでいない。漿膜があるために卵の外部からはみえないが、内部では胚の頭部表面がキチン化し、褐色を呈する。臍孔は完全に閉鎖。固定した胚を色素で染めると、胴部が未だよく染まる。ほぼ水野の己Cに当るものと思われる(第25図f)。不越年性卵の7日半。気管にガスが入る(637)。
Stage 27 点青Ⅱ期 胚が漿膜を飲み込み、着色した頭部は卵殻を通して青みを帯びてみえる。点青または目付きと呼ばれる時期で、孵化の前々日に当る。胴部は未だ着色が極く淡いが、色素に染まり難い。孵化を揃えるために、この時期から光を遮断して催青するのが普通である。不越年性卵の8日目。
Stage 28 催青Ⅰ期 胚は頭部ばかりでなく胴部も着色するため、卵全体が青みを帯びた色にみえる。催青卵または青み卵と呼ばれる時期であるが、胴部の着色が未だ十分に濃くないため、生卵をそのままみると、頭部の色が濃く、胴部との識別が容易である。この段階は点青後期と呼ばれていることもある(986)。不越年卵では9日目。
Stage 29 催青Ⅱ期 胴部の着色が濃くなり、生体のままの卵の外観では、卵全体が一様に青み、頭部をはっきりと識別し難い。孵化直前の状態である。不越年性卵は9日半でこの時期に達する。催青卵の卵殻を切破って胚を出し、乾燥を防いで飼育すると、自然に孵化したものと同様に発育し得る。室賀(613)は欧19号の青み卵になったばかりの卵から胚を取出して発育させることができたと云っているが、大村・菅生(757)によれば、孵化前日20時の胚は取出しても正常と大差なく育ち、同12時の胚は表皮柔かく扱いにくいが15%が上蔟し、同3時の胚は柔かく水々しているが3%が3令に達した(以後の飼育中止)。孵化前々日9時の胚は白く水々しているが、4%が3令に達した。
Stage 30 孵化期 孵化。不越年性卵では10日目。孵化の行動については室賀(607)、森山(595)、中島(653)、竹内(983,984)らの観察がある。
不越年性卵や人工孵化種の胚は、Stage7 から直接Stage14に眺んだような形態変化を極く短時間のうちに経過し、卵黄細胞の移動も漿膜の着色も起こらない。ただ不越年性卵が越年性卵と異なるのは、胚に18体節の全部が現われないうちに、頭胸部になる部分の体節構造が特に明瞭になり、正中線も認められることである(129,450,668)。産卵後約10日で孵化するが、器官形成期に入ってからの経過は、時間的にも形態的にも、越年性卵と大差がない(第154-156表)。
3 生理
蚕種の取扱いに関係の深い問題だけについて述べる。
A 水分
産卵当時の蚕卵は平均65%ぐらいの水分を含んでいるが、日のたつに連れてその絶対量が減少し、6月採種の越年性卵は翌年3月までに約6%の水分を失なう(671)。しかし、この間に固形分も4-5%減少するので、水分率としては殆ど変化がないとされている。勿論これは保護条件によって相違し、湿度の低い場合には水分の減少が大きい(Ⅵ3Db)。催青に着手すると蚕卵重量の減少は著しく、孵化前日までに約10%減るが、水分率はほぼ66%を保持している(367)。蚕卵は、卵殻の構造上、長時間水に浸しておいても内部へ水を通さず、水分率に殆ど変化がない(783)。
このように、程度の差はあっても、水分は産卵後減少の一路を辿り、タト部から補給されることはないのであるが、卵殻の着いたままの卵全体ではなく、毛細管で採種した卵の内容だけについて測定すると、胚発育の時期に応じて著しい水分率の増加または減少が認められると云う(466,467)。黒田らはこの水分微量測定法によって調べた結果、外部からの吸水の考えれない蚕卵の中で、産卵後3日間に6%以上も水分率の増加していることに注目し、この増加した水は、胚の成長に伴なって貯蔵養物が“酸化分解され又縮合される等新陳代謝の結果生ずる”代謝水であると考えた(467)。
梅谷(1101,1106)は、この方法によって調べると、越年卵においては、1)産卵後2日目(ダルマ期)、2)7-10日目(アレイ期)、3)越冬後、胚が発育を始めて付属肢突起を形成する時期、および4)反転後に水分率の低下がみられるが(第26図)、これらは何れも代謝作用の緩慢な時期であって、低温に耐える力も強いとし、広く卵態越冬の昆虫をみると、総べて蚕のこれらの時期の何れかに相当する段階で越冬しており、蚕の胚は2)の時期で越冬すると論じている。但し、この解釈はバッタの卵の生理作用の消長と一致しない点があるが、それは測定法の相違によるのではないかと云う。伊藤(278)は、催青中の卵において、梅谷が抵抗力の弱いと云っている時期にほぼ該当する反転前乃至反転期および孵化前に尿酸量の減少することを認め、抵抗力との平行関係に注目しているが、生理的な意義は不明で、平行現象として指摘したに過ぎない。
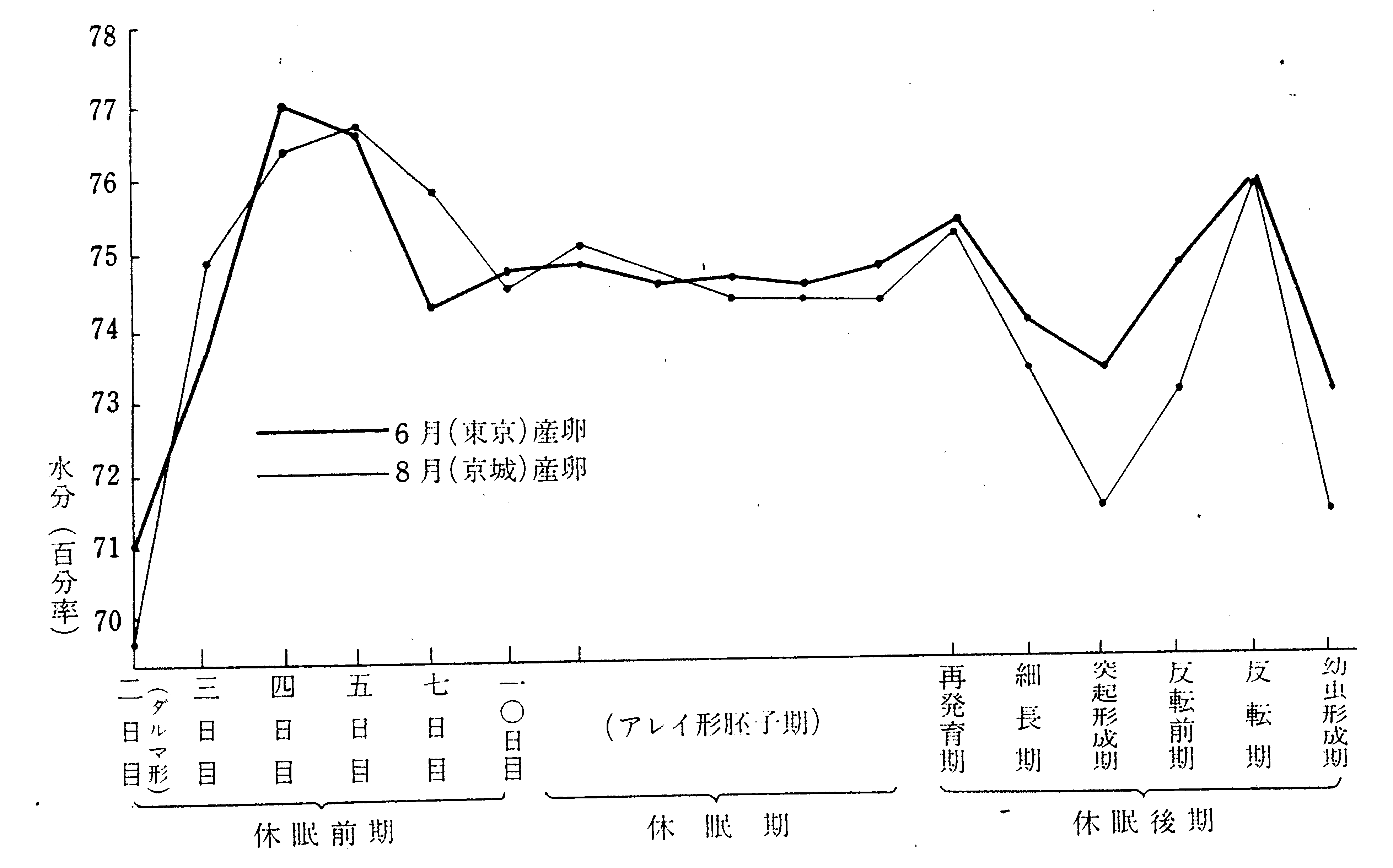
第26図 越年性卵における水分量の時期的変化(梅谷)(1106)
越年性卵においては、休眠が進むと卵黄細胞が周辺部に移勤し、卵の中央部に液質の部分ができ、冬中この状態が続くのであるが、これが水分微量測定の値にどのように影響するかについては言及されていない。しかしこの卵黄細胞の偏在は凍結実験の結果には次ぎのように影響している。
青木(46)によれば、越年性卵を-36℃ぐらいの低温で凍結させて凍結曲線を調べると、-15~-20℃で過冷却が破れて凍り初め、温度は一且-5℃付近まで急上昇するが、再び徐々に下がる。この凍結曲線の下降部は産卵後1日目には滑かで、凍結点は一つだけであるが、3-4日目頃から下降曲線の-13℃付近に第2凍結点が現われ初め、10日目には極めて明瞭な平坦部として認められる。このような凍結曲線の形は産卵後100日間も続くが、多価アルコールの存在(Ⅴ2B)とは関係がない。凍結した卵を速かに融かして再凍結を行なうと、1度目に完全に凍結させた卵には第2凍結点が現われないが、1度目の凍結を第2凍結点の現われる前に止めて融かした卵にはこれが現われること、未受精卵の凍結曲線にも第2凍結点は現われないこと、などから考えると、休眠の進行に伴なって卵内に凍り易い部分と凍りにくい部分とのできることが考えられる。これは卵黄細胞の移動によって卵の中央部(この部分に熱電対を挿入して測定した)に液質の部分のできることとよく平行している。越冬後、丙B期頃に達した卵においては第2凍結点が現われないとも聞いている。
越年性卵にグリセロールおよびソルビトールの増加するのは冬期間の凍結防止に役立っているのではないか、と云う考えはこの凍結実験の結果によって否定されたが、蚕卵は休眠状態で冬を越すものではなく、これらの多価アルコールの減少し初めたか、もしくは減少してしまった状態で厳冬期を過すことからみても、その生態的意味を大きく考えることはできない。
蚕卵からの水分蒸散は、即時浸酸種についての測定によれば(698)、25℃、湿度75%の場合に、浸酸後3時間半で最高(1.470mg/g.hr)を示し、以後急激に減少し、翌日には0.551、3日目には0.519で最低に達する。その後は次第に増加し、9日目(孵化前日)には初日にほぼ等しい価(1.313)に回復する。この変化は梅谷の再発育期以後の代謝水の増減(第26図)と傾向を同じくしていると云う。
休眠に伴なって胚および卵黄細胞から水が出て、それらの組識は収縮しているのであろうと云うことを先きに述べたが、この時期の蛋自性卵黄顆粒には中性赤による顆粒状染色(コアセルヴェーショソと考えられる)が起こりにくいことからみて、蛋白質分子の水和性が強くなっているのではないかと云う想像もできる(929,948)。
B 呼吸
a 不越年性卵
催青中の毎日の酸素消費量(105,364,680,681,683,684)、同じく炭酸ガス排出量(368,893)などには多くの研究がある。
不越年性卵および浸酸卵の炭酸ガス排出量または酸素消費量は、催青初期には日々の増加が明瞭でないが、反転後から増加し、点青期頃に急増する(第97表)。
| 卵 令 | 不越年種の炭酸ガス排出量 (五大州、26℃)(g) |
即時浸酸種の酸素消費量 (日111号、25℃)(g) |
| 第2日 3 4 5 6 7 8 9 10 |
8.7417 10.0207 12.3540 8.4248 9.9410 14.8400 21.9980 31.4351 32.2519(一部孵化) |
8.072 7.248 9.083 7.438 9.514 12.065 13.803 16.605 31.977(催青卵) |
これらの排出量および消費量は温度によって著しく変化するが、西郷(781)によれば、点青期の酸素消費量と温度との関係はArrheniusの式
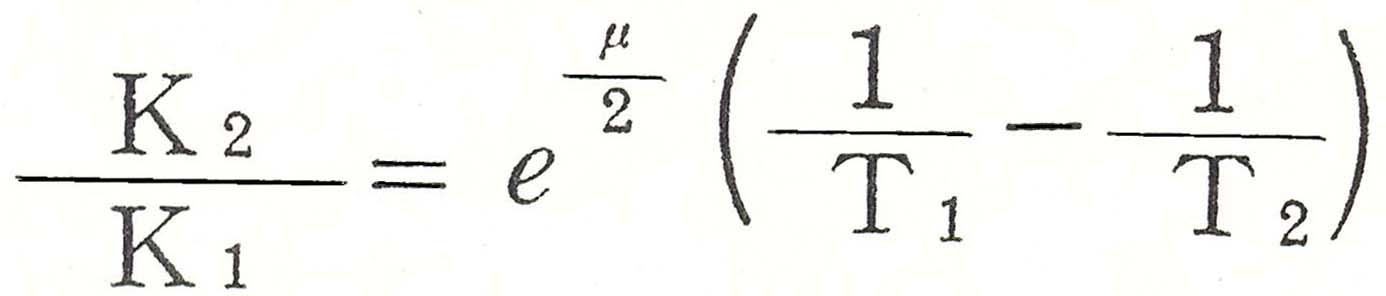
によって表わされ、μの値は5-15℃では25,000、15-25℃では15,700、25-37℃では6,000であり、Q10は5-15℃で4、15-25℃で2.5、25-35℃では1.5であると云う。
川瀬(364)は、催青中のチトクロームcオキシダーゼ作用の増大が7日目(9日目に1部が孵化)までは顕著でないがその後急増することと、卵の酸素消費量が7日目以後に著しく増加することとの間には関係があるものと考えている。
入戸野・中曾根(684)は、休眠中の卵の酸素消費量は酸素分圧によって著しく影響を受けるがCOによる阻害を受けないこと、休眠を終えると酸素分圧の影響が減じるがCOによる阻害が現われ、殊に催青末期のCOによる阻害の著しいこと、COによる阻害は光線の有無によって影響されることなどから、休眠中の呼吸にはフラポプロテイン系酵素が、休眠を終えた催青中の卵の呼吸においてはチトクローム酸化酵素が、それぞれ末端酵素として働いているのであろうと考えた。
西沢ら(681)によれば、反転直後2日間ほどは青酸による酸素消費の阻害が特に著しい。
山ロ・清水(1165)は、卵そのまま、卵の磨砕物、胚だけを取出したもの、および胚の磨砕物の酸素消費量を比較し、催青中の胚においては、呼吸作用の70-40%は胚の“構造”に帰すべき部分、30-60%は酵素その他の、細胞の基礎的な生化学的組成に帰すべき部分であろうと云っているが、詳細は不明である。
催青中を通じて、呼吸商が0.6-0.7であることから(368,683)、そのエネルギー源は主として脂肪であろうと考えられている。
催青中の酸素分圧は170mmHg内外が最適で、200または140mmHgでは催青死卵が出ると云う(604)。
催青中の所要空気量あるいは換気回数などを決定する基礎資料としては、前記鈴木の炭酸ガス排出量が用いられている。重量で出ている排出量を体積に換算し、その量だけの炭酸ガスを許容量まで稀釈するにはどれだけの空気量が必要かと云う計算をするのであるが、これには気圧、温度、空気中の炭酸ガス量、炭酸ガス許容量などを測定しなければならない。
簡便法としては、炭酸ガス許容量(P)を1%(1㍑中10cc)、空気中の炭酸ガス量(Q)を0.04%(1㍑中0.4cc)とし、気圧は無視して計算する方法が用いられている(605,606)。体積に換算した炭酸ガス総排出量をK(cc)とすれば、所要空気量(Y)は
![]()
である。室賀(620)は催青中の所要空気量を、この方法によって蚕種10g当り88.2㍑と計算した。許容量を0.5%にとった鈴木(884)の計算では178㍑となっている。
しかし、これらの値は、蚕種を実際に催青した結果から考えられる所要空気量に比べて、非常に大きな空気量である。室賀自身も、毎日1回換気すれば、10gの蚕種に対して1日2㍑でほぼ足りると云い、三浦(547)は、40,000粒の蚕種の催青には、1㍑の容器に収容して1日2-3回換気すればよいと記載している。古い横田ら(1194)の成績によれば、容績1㍑のガラス器内に密封して、全期開催青しても害のない蚕種(不越年)の量は2-3蛾(1蛾400粒内外)以内であるが、1㍑に15-16蛾分を密閉しても、催青の前半だけならば被害がないと云う。杉山・高見(877)のガラス器に密封した試験結果では、10gの蚕種は全催青期間を通じ20㍑の空気があれば十分に孵化した。1日2㍑の空気量では、催青初期には十分であるが、末期にはやや不十分であった。芦田(68)は、即時浸酸種1gを30日間冷蔵(5℃)する所要空気量を20cc以上としている。
人工孵化種を催青初期に輸送する場合には、輸送に2日かかるものとすれば、蚕種10g当り多めにみて12㍑の空気量があれば、密閉して送っても呼吸に支障がないと云う計算(591)が示されている。これは上記の方法によって計算したものであると云うが、実際問題としては、このように大きな容積の荷造りをして蚕種を送ることはない。横田ら(1194)によれば、木箱であれば、越年種、不越年種ともに1㍑に100蛾分を密封しても催青初期8日間は被害がなく、この倍量を入れても初期5日間以内ならば差支えなかった(温度は75゚F前後)。
このような計算と実際との不一致を、室賀(622)は、計算量は呼吸障害を蒙るような空気には絶対に接触させない場合を考えた量であり、実験例として報告されているのは、呼吸障害を蒙るような不良な空気に接触しても、その期間が短かいために害がなかった場合であろうと考えているが、なお検計すべき問題である。
卵殻だけを除去した卵内容全体を液体培地で培養すると、点青期まではよく発育して、漿膜も嚥下するが、ここで気管系を通じての呼吸に切り変わるため(637)、胚は溺死する(941)。
b 越年性卵
越年性卵が休眠に入ると呼吸量が著しく減少するが、これは単なる量の減少ではなく、青酸やCOによる阻害の減少から考えてもわかるように質的な変化である。越年性卵における酸素消費の青酸阻害は産卵後2日目頃までが最も著しく、1.5×10-4Mで阻害が現われるが、4日目以後では3×10-4Mになり、以後90日になってもこの状態が続くと云う(785)。
休眠に伴なう糖代謝の機構を追究した茅野(Ⅴ2B)の研究は、休眠卵の呼吸について注目すべき結果を示している。グリコーゲンからのソルビトールおよびグリセロールの生成反応には多量のNADH2およびNADPH2が必要である。不越年性卵においては、これらの補酵素はミトコンドリアの呼吸酵素系によって速かに酸化されるが、越年性卵においては、正常の呼吸酵素系が何処かの点で遮断されるために、これらの補酵素の酸化が止まり、多価アルコールの生成に向けられるようになる。休眠に入ってからの脂質の減少量は、低下した酸素消費量(第27図)によっては到底CO2およびH2Oに完全に酸化し得ないほど多量であるから、脂質からの水素はそのまま酸素に行くのではなく、何らかの中間代謝物に移されているものと考えなければならないが、これは呼吸商の増大からも推定される。

第27図 休眠初期の蚕卵における酸素消費量および呼吸商(Chino)(82)
○:酸素消費量、×:呼吸商。測定は25℃。
NADH2およびNADPH2は解糖、ペントース燐酸径路およびリンゴ酸酵素によるリンゴ酸塩の酸化などの過程で作られて多価アルコールの生成に用いられ、この生成は無酸素状態でも阻害されない。しかし、休眠を終えて多価アルコールからグリコーゲンの再合成される反応には酸素が必要である。
更に特異なのは、蚕卵ホモジネートの吸収スペクトルによって確認できるチトクロームはb5であり、cは、孵化の3日ぐらい前になって微量に存在するのではないかと考えられる程度の量しか見出されないことである。チトクロームcオキシダーゼの存在はスペクトルで確認できる。b5はミトコンドリア分劃には極く僅かで、主として、脂質に富む分劃に含まれており、この分劃にはチトクロームcオキシダーゼも多い。チトクロームb5およびチトクロームcオキシダーゼの最大吸収部は、休眠に入ると次第に消失し、産卵後10日ぐらいで殆ど認められなくなる。休眠から醒めると再び現われるが、休眠初期における多価アルコールの出現はこれらの消失よりも早く、また休眠から醒めると低温下でもグリコーゲン量は回復するが、チトクロームb5およびチトクロームcオキシダーゼは催青を始めなければ出現しないから、チトクロームの消失が多能アルコールの生成の第一義的な要因とは考えられないと云う。
茅野は実験の結果を綜合して、前休眠期および休眠後の催青初期には、NADH2またはNADPH2からフラボプロテイン、チトクロームb5、チトクロームc、チトクロームcオキシダーゼを通る特殊な径路で酸素への電子伝達が行なわれ、催青末期に筋肉が発生するようになるとミトコンドリア分劃にチトクローム酵素が集中して、普通の電子伝達系が働くようになるのではないかと考えている。石川(258)は休眠に伴なう上記補酵素の酸化型と還元型との消長を調べている。
越年卵を冬季間、自然温度の水に浸漬しておくと、胚に付属肢の形成が始まる前までは、発育はややおくれるが自然温度で保護した無浸漬卵に比べて孵化歩合は劣らず、寧ろ高い場合があるが(782)、付属肢形成が始まってからも浸漬を続けると、急激に孵化歩合が低下すると云う。また、産卵後3カ月ぐらい高温で保護した卵を5℃の水に浸すと230-250日の浸漬に耐え、高温30-60日の卵は同じく140-170日の浸漬に耐えたと報告されているが、著者らの試験(末発表)では、産卵後、25℃、30日で浸水した場合に、孵化歩合が50%に低下する浸漬日数はこれに比べて非常に短かく、5℃で30日、25℃で3日に過ぎなかった。その原因は明かでない。著者らの場合には、水の腐敗を防ぐために卵を消毒し、滅菌蒸溜水に浸漬した。
C 発育
a 胚の成長
熱湯で固定した越年卵から胚を取出し、その乾物量を測定した山口ら(1154,1162)の結果によれば、
催青期間の10-20%を経過した最長期乃至突起発生期
〃 45% 〃 反転期
〃 70% 〃 気管形成期あるいは口器着色期
をそれぞれ中心とする成長率の高い時期があり、30%(反転前)および50-60%(反転後)を中心にする成長率の低い時期がある。木下(380)もほぼ同様な結果を報告している。
山口らはこの成長率の変化は梅谷の考え(Ⅵ3A)によく一致すると云うが、別の見方をすれば、最長期乃至突起発生期は盛んな細胞分裂と共に器官形成の始まる時期であるから成長率の高いのは当然であろうが、反転期を中心とする時期の重量成長には、背面形成によって体内に包み込む卵黄の重量がみかけの成長として加算されているのではなかろうか。また、気管形成期あるいは口器着色期と云うのが、木下の報告のように点青期のことであるとすると、漿膜を飲み込んだための体重増加が関係しているかも知れない。これらの点を吟味した上で検討する必要がある。
| 測定月日 | 体 長 (μ) |
体 巾 | 胚の呼び名 | ||
| 頭葉(μ) | 胸部(μ) | 尾節(μ) | |||
| 1月15日 30 2月 5日 25 3月 5日 10 15 20 25 30 4月 5日 10 |
1,305 1,286 1,165 1,252 1,332 1,431 1,403 1,564 1,435 1,546 1,454 1,779 |
344 320 315 350 347 351 352 393 351 347 338 399 |
220 198 202 229 207 220 203 246 207 214 199 210 |
225 192 196 204 187 181 194 213 189 218 194 216 |
┐ │ ├水野甲、梅谷A-B │ ┘ ┐ ┴水野乙A-乙B、梅谷C ┐ │ ├水野丙A、梅谷D ┘ 水野丙B、梅谷E(細長期) |
| 固定時期 | 体長 (mm) |
体 巾(mm) | 胚の発育階程 | |||
| 第4環節 | 第6環節 | 第8環節 | 第13環節付近 | |||
| 1月15日 30 2月15日 28 3月15日 30 |
1.45 1.53 1.75 1.75 2.11 2.19 |
0.19 0.19 0.21 0.23 0.20 0.22 |
0.20 0.20 0.24 0.25 0.21 0.23 |
0.17 0.19 0.21 0.22 0.19 0.21 |
0.17 0.18 0.21 0.21 0.18 0.20 |
甲 乙A+ 乙B- 乙B+、丙A 丙B 丙B+ |
b 卵重の変化
蚕卵の重量は産卵後の経過に連れて減少する(第100表)。
| 蚕品種 | 蚕種の種類 | 基準卵重 | 催 青 | 孵化 | 毛蚕 | |||||||||
| 直 前 | 第1日 | 第2日 | 第3日 | 第4日 | 第5日 | 第6日 | 第7日 | 第8日 | 第9日 | |||||
日122号×支115号 支115号×日122号 |
春採越年 〃 |
g 0.2772 0.2822 |
% 8.0 3.9 |
% 8.6 4.2 |
% 8.8 4.4 |
% 9.1 4.5 |
% 9.4 4.8 |
% 10.3 5.5 |
% 11.4 6.6 |
% 12.8 8.3 |
% 14.6 10.9 |
% 16.4 ─→ |
% 18.1 12.5 |
% 29.1 28.7 |
|
日122号×支115号 支115号×日122号 |
秋採越年 〃 |
0.2637 0.2629 |
3.8 2.4 |
4.1 2.6 |
4.2 2.7 |
4.4 3.0 |
4.9 3.3 |
5.7 4.1 |
7.1 5.4 |
8.9 7.2 |
11.4 9.9 |
─→ ─→ |
14.2 11.3 |
27.6 26.2 |
|
日122号×支115号 支115号×日122号 |
春採即浸 〃 |
0.2953 0.2886 |
─→ ─→ |
1.2 1.2 |
2.2 2.2 |
2.8 3.3 |
3.9 4.4 |
5.2 5.5 |
6.6 8.0 |
10.2 ── |
── ── |
─→ ─→ |
12.2 10.3 |
26.2 25.6 |
|
日122号×支115号 支115号×日122号 |
春採即浸冷 〃 |
0.2981 0.2890 |
─→ ─→ |
2.9 3.6 |
4.0 4.9 |
5.2 6.2 |
6.3 7.6 |
8.5 10.3 |
11.8 ── |
── ── |
── ── |
─→ ─→ |
14.3 13.3 |
26.8 29.5 |
|
日122号×支115号 支115号×日122号 |
春採冷浸 〃 |
0.2965 0.2952 |
─→ ─→ |
0.8 0.6 |
1.3 0.9 |
1.7 1.2 |
2.4 1.7 |
3.3 2.6 |
4.7 3.7 |
8.0 6.2 |
── 9.1 |
─→ ─→ |
11.4 11.0 |
28.4 28.3 |
山口(1158)は、催青中の卵重量の減少は直線的ではなく、初期はやや緩慢であり、催青期間の50-60%を過ぎると急降下すると云い、水分率や卵汁の理学的性状(浸透圧、電気伝導度、比重、粘性)などの変化をも調べた結果を綜合して、卵の発育には反転期頃を分岐点とする生理的二期があるとし、反転期は生化学的作用が旺盛であると云う梅谷の説(Ⅵ3A)とはいささか矛盾すると考えている。この場合、反転期の中心は、24℃内外の催青において、丙Bから蟻蚕完成(孵化の前日)までの経過の大約45%の点であった。
鈴木の測定した不越年性卵の炭酸ガス排出量(第97表)をみると、産卵後5日目頃に一時的な排出量の減少が認められるが、徳永(1056)はこの現象に注目し、鈴木の成績においては五大洲の卵期間の44%、青熟(表には省略)の同40%の時期にこの排出量の低下していること、また水野ら(574)の調査した即時浸酸種の減耗量についても、日本種春採り3品種平均卵期間の41.7%、同秋採り3品種平均および日支交雑種2品種平均のそれぞれ卵期間44.4%の時期に減耗量割合が一時的に低下していることを指摘し、不越年性卵の成長にも越年性卵にみられる休眠に類似の現象があり、そのために二期的な成長をするのではないかと考えている。しかし、炭酸ガス排出量については兎も角として(川瀬ら(368)の4日目の低下は鈴木の場合のように顕著ではない)、卵重量の減耗は、山ロの場合、水野らの場合および第100表の成績がそれぞれに異なるようであるから、なお検討の余地があるように思われる。即浸卵の4日目頃に減耗量の低下するのは、布目らのみている蒸散量の低下(Ⅵ3A)と一致するのかも知れない。
D 環境
a 温度
催青中の胚発育は温度によって著しく影響を受ける(第101表)。
| 蚕 品 種 | 20℃区(%) | 24℃区(%) | 28℃区 |
| 日 1 号 支 4 号 支 7 号 支 9 号 欧 7 号 欧 9 号 日107号 正 白 支101号 新 白 |
89 92 84 81 82 88 94 90 95 96 |
90 89 90 93 86 90 93 91 95 95 |
85 93 83 93 69 88 89 84 93 95 |
松村・石坂(515)が第101表と同様な3回の試験結果を平均した孵化歩合は、20℃区92.54%、24℃区93.02%、28℃区88.88%であったが、20℃区は24℃区または28℃区に比べて孵化の不斉な場合が多かった。
10品種平均の胚発育速度(催青日数の逆数)は、湿度72%場合に、20℃区0.064、24℃区0.090、28℃区0.106であった。孵化した蟻蚕の体重は28℃区が最も軽く、絶食生命時間は 24℃区>20℃区>28℃区であった。
赤卵や白卵の越年種には孵化の悪いものが多いが、これらは夏期の保護温度および催青温度を25℃よりも低目にすると孵化がよくなる(964,1115)。
これらの結果を綜合すると、胚の発育に最適な催青温度は20℃以上25℃以下であることがわかる。
なお蟻蚕の体色は催青温度の高いほど淡く、低いほど黒味を帯びる。蟻蚕の体色に温度の影響するのは胚の形態が完成し、皮膚の着色する時期である(1135)。松村(512)および長島(635)はメラニン色素の形成を調べ、温度と蟻蚕体色との関係を化学的にも証明した。松村は、産卵直後の保護温度が高いと卵色の淡いこと(Ⅴ2A)をも同様にメラニン色素の濃淡によるものとしているが、これは主として3・ヒドロキシキヌレニンの色素形成のための消費量が温度によって相違するためであろう。蚕卵においてはメラニン系色素は検出できないと云われている(717)。褐円斑紋の発現は、反転期の催青温度が25℃以下の間は温度の上昇に連れて低下するが、25℃以上になると反対に高まるらしい。即ち10℃≧15℃>20℃>25℃<30℃<35℃であった。
越年性卵の漿膜細胞色素粒は、産卵後、高温の続く間は細胞内に一様に分布しており、そのために卵色は濃くみえるが、気温が低下すると核が細胞の一端に偏在するようになり、色素粒もその周辺に集まってしまうので、卵色が淡くみえるようになる。冬が過ぎて気温が上ると色素粒は再び細胞内に一様に分散する(533,534,670,1137)。自然温度で蚕種の冬期保護を行なう場合に、漿膜色素の分散状態が卵の活性比状態を知る一つの指標になるように云われることがあるが(954)、特定な品種を特定な地方で毎年みているような場合は別として、一般的には、色素数の移動と卵の活性化状態とは必らずしも一致せず、色素粒の集積したときに既に活性化のかなり進んでいるものもあれば、全く活性化していないものもある。胚の形とこの移動との間にも直接の関係はない(1087)。
休眼中の温度については既に述べた(Ⅴ1B)。
休眠を終えた胚が孵化まで発育し得る最低温度は12.5℃であるが、点青期以後ならば10℃、催青卵ならば7.5℃でも孵化し得ると云われている(905)。若い胚ほど低温においてもよく発育するのに(第103表)このような結果になるのは、低温においては発育に非常な長期間を要するために、若い胚から初めると途中で活力が低下してしまい、孵化にまで達し得ないのであろうと考えられているが、臨界期までの発育は胚の伸長に過ぎず(Ⅵ2B)、物質合成を必要とする器官形成期の成長は低温では十分に進行し得ないのであろうと云うこと(702)も考える必要がある。
胚の長さの伸長を発育とみるならば、休眠を終えたばかりの胚は0℃においても僅かながら発育する(第28図)。
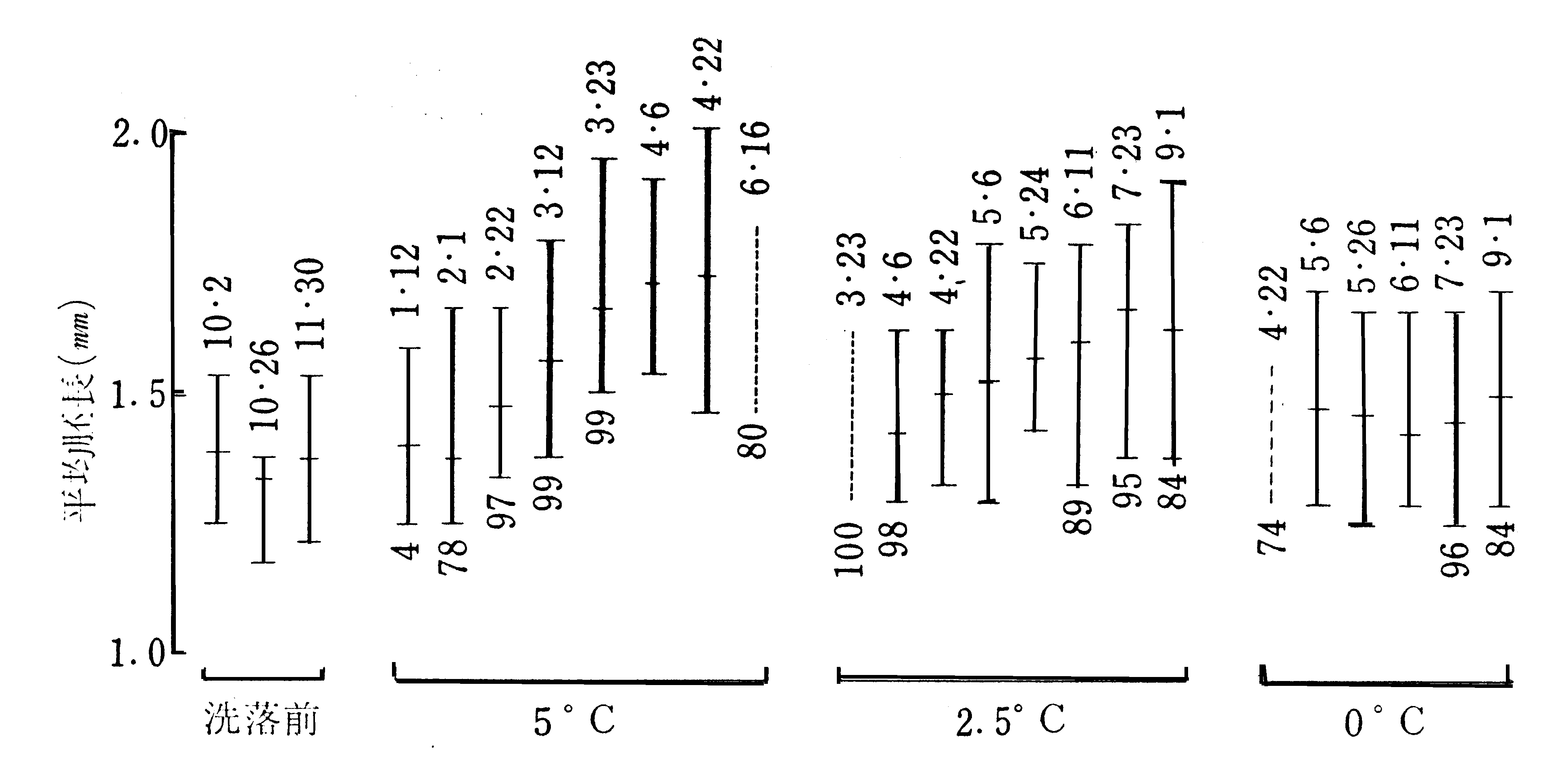
第28図 冷蔵温度別の胚発育(高見・戸谷ら)(945)
日124×支124のF2卵(晩秋採り)。12月3日に洗落し5℃、2.5℃および0℃に分けて冷蔵した。
最上段の数字は月日で、その時測定した胚の長さの分布を縦線の長さで表わす。短かい横線は平均値。
核分裂の認められなかった場合は細い縦線、認められた場合は太い縦線で示した。縦線の下の数字は
催青2週間孵化歩合。点線の場合には胚長は測定しなかったが、孵化歩合は調べた。
この図において注目されるのは、活性化、胚の発育および核分裂はそれぞれに必要な最低温度を異にしていることである。
0℃においては活性比はするが、胚の発育は僅かに進むだけであり、核分裂は全然起こらない。
2.5℃におては、活性化および胚の発育(丙Bまで)は進むが、調査した期間内では核分裂は超らなかった。5℃においては、活性化、胚発育および核分裂が共に進行する。即ち、臨界期までの発育は伸長を主とするものであり、物質合成を伴なう真の成長はその後に始まると考えられる。
この点に関連して興味があるのは鈴木(893)の炭酸ガス排出量に関する試験である。9月3日産卵の青熟越年卵を2月12日から151日間、+5℃、-2.5℃および-5℃に保護して、その間の炭酸ガス排出量を測定した結果から蚕卵1kg当りの1日量を計算すると、+5℃ 0.28660g、-2.5℃ 0.00379g
、-5℃ 0.00322gで、-2.5℃と-5℃との差は小さいが、+5℃と-5℃との間には生理的に大きな断層のあることがわかる。
水野によれば、冷蔵中の胚発育は第102表、胚の階程別発育速度は第103表の通りである。
| 冷蔵当時 の胚 |
冷蔵温度 (℃) |
冷蔵日数 | ||||||||
| 20日 | 40日 | 60日 | 80日 | 100日 | 120日 | 140日 | 160日 | 200日 | ||
| 甲 | -2.5 0 2.5 5 7.5 |
甲 甲 甲 (-)乙B (-)丙A |
甲 甲 甲 (+)乙B 丁B |
甲 甲 甲 (-)丙A (+)戊 |
甲 甲 甲 丙A 戊B |
甲 甲 乙B 丙B (+)戊B |
甲 (+)甲 乙B (-)丁A (+)戊B |
甲 (+)甲 乙B (-)丁A (+)戊B |
甲 乙A 乙B (-)丁A - |
甲 - - - - |
| 丙B | 0 2.5 5 7.5 |
丙B 丙B 丙B (-)丁B |
丙B (+)丙B (+)丙B 丁B |
丙B (+)丙B 丁A 戊B |
丙B (+)丙B (+)丁A (-)戊C |
丙B (+)丙B (-)丁B (-)戊C |
丙B (+)丙B (-)丁B (-)戊C |
丙B (+)丙B (-)丁B (-)戊C |
丙B (+)丙B (-)丁B - |
- - - - |
| 目的の 発育期間 |
保護温度 (℃) |
発育所要日数 (日・時間) |
積算温度 (時間) |
目的の発生段階に達した胚の割合 |
| A 休眠期(甲) から 最長期(丙B) |
30.0 27.5 25.0 22.5 20.0 17.5 15.0 12.5 10.0 |
3・12 3・12 3・12 3・12 3・20 4・10 4・17 6・10 11・03 |
2,520 2,310 2,100 1,890 1,840 1,855 1,695 1,925 2,670 |
5%。他は総べて若く、極めて不斉。 8%。他は大部分若く、極めて不斉。 24%。 〃 〃 44%。不斉。 55%。やや不斉。 90%。斉一。 92%。 〃 90%。 〃 91%。 〃 |
| B 最長期(丙B) から 反転期 |
30.0 27.5 25.0 22.5 20.0 17.5 15.0 12.5 10.0 |
3・14 3・14 3・14 4・02 5・02 7・13 9・19 14・16 33・17 |
2,150 1,935 1,720 1,715 2,130 2,263 2,350 2,640 4,045 |
全部。斉。 〃 〃 〃 〃 〃 〃 93%。他は若く、ほぼ斉一。 75%。他は老幼やや不斉。 67%がほぼ反転、他は老幼やや不斉。 52% 〃 他は老幼不斉。 57% 〃 〃 |
| C 反転期 から 催青期 |
30.0 27.5 25.0 22.5 20.0 17.5 15.0 12.5 10.0 |
4・06 4・10 4・18 5・18 7・16 11・20 16・00 30・00 55・00 |
2,550 2,385 2,280 2,415 2,760 3,550 3,840 5,400 6,600 |
斉一。 〃 〃 〃 〃 〃 〃 〃 〃 |
第103表において注目されるのは、Aのように若い区間の発育所要日数は、30℃でも15℃でも著しい相違がないが、Cのように進んだ区間の発育所要日数は温度が低いと非常に長いことである。これを逆に云えば、胚の発育程度の揃っていない蚕種を高温で催青すると発育の開きがますます大きくなるが、低温で催青すると、進んだ階程の胚の発育は比較的おそくなり、おくれた階程のものの発育が比較的進んで、全体としての発育の開きが小さくなる。これは蚕種の取扱い上実際に利用される重要なことがらである。
低温のために発育を抑えられていた胚は、高温に移すと発育が非常に速い。例えば、第28図の場合に、2.5℃区4月6日の胚は平均体長1.43mmであったが、これを10℃に移して2日おいたものは1.58mm、15℃1日では1.55mmに発育しただけで核分裂は認められなかった。これに対し、25℃に移したものは1日で2.01mmとなり全部の個体(11頭)に核分裂が認められた。
蚕卵の耐え得る低温の限度についての奥村(707)の試験によれば、越年卵を2月に-20℃の恒温に冷蔵すると、2日間でも著しく孵化歩合が低下したが、北海道の冬季間のように、-20℃以下の日はあっても、間歇的な場合には被害がなかった。仲野(656)によれば、休眠期胚子(甲胚子の意味)は-5℃に14日接触させても孵化が斉一であったが、-10℃14日間または-15℃3日間では孵化不斉になった。
丙B以後、孵化前日までの催青中の卵の耐寒性について、日9号×支108号、日114号×支114号および欧18号×支107号を用いて行なった山口(1156)の実験によれば(第104表)、催青期間12日の場合の5日目(経過歩合約42%、反転期)頃において、その前後に比べて耐寒性の弱い時があるように思われると云う。なお、この場合の不発生卵は大部分が催青死卵で、普通死卵は1-5%(10%を越える例は極めて少ない)であったから、この程度の低温は胚に対して直接の致死作用をおよぼすものではないと考えられる。
| 催青日次 | 低温接触 | |||
| -10℃、6時間 | -20℃、6時間 | -10℃、24時間 | -20℃、24時間 | |
| 第 1日目 2 3 4 5 6 7 8 9 10 11 12 |
90% 81 83 75 64 80 74 81 67 14 10 0 |
74% 82 75 67 63 73 40 24 6 0 0 0 |
79% 82 72 80 78 80 70 23 2 0 0 0 |
78% 67 71 57 29 12 14 2 0 0 0 0 |
先きに述べた青木の凍結実験(Ⅵ3A)においては、-15℃~-20℃で凍結が始まったが、これは熱電対の挿入によってにじみ出た卵黄に氷ができ易いためで、自然状態で凍結の始まる温度はこれよりも低いと云う。
梅谷(1114,1116,1124)は交尾した雌蛾の産んだ卵を受精前に5℃と2.5℃とに冷蔵した処、5℃のものは60日間冷蔵した後にも70%以上の受精率を示したが、2.5℃においては1日間の冷蔵によっても受精率が30%ぐらいに低下したと云い、これは卵核ではなく精核が被害を受けるのであって、2.5℃と5℃との間に精子生存の臨界点があると考えた。これに対し、橋本(183)は、産卵後5-20分の卵を2.5℃、0℃、-2.5℃にそれぞれ42-43時間冷蔵した後に、高温処理を施して雄核発生(Ⅵ2A)を行なわせた結果から、産卵直後の低温に対しては精核よりも卵核が弱いと報告している。
滝沢・玉沢(1012)は、交尾蛾の産んだ1時間以内の蚕卵を冷蔵すると、-20℃においては2日間、-15℃においては10日間、-5℃および0℃では60日間ぐらいで受精力を失なうが、5℃においては60日間でも受精力を失なわず、しかも40日前後で淡褐色に着色し初め、受精卵と不受精卵との区別ができるようになるから、5℃では受精が行なわれると云っている。また、蛾を過冷却すると雄蛾よりも雌蛾の受精力の減退が著しいから、梅谷の考えには同意し得ないとしている。交尾蛾の卵を産卵後30分以内に冷蔵した結果からみると、-5℃と-10℃との間に抵抗力の臨界点があるらしい(1011)。
産卵後1-2時間目の卵が産卵直後の卵よりも低温(0℃)に対する抵抗力の弱いのは(618)、この時期に第二成熟分裂および受精の行なわれるためと考えられ、低温ばかりでなく、種々な障害に対して弱い時期である。山ロ(1155)は、産卵後25℃で2時間目を蚕卵標準抵抗力曲線の第1降下点と呼んでいる。第2降下点は産卵後12時間目の、胚盤から胚帯の形成され初める時で、人工孵化による畸形が生じ易い。20時間目以後は障害に強くなる。
蚕卵に対する低温の影響については、上記のように、条件の設定と受精または孵化とを直接に結び付けた報告が大部分で、その過程の分析がないために、結果の相違があってもその原因を究明し得ない場合が多いが、冷蔵によって卵内にある種のアミノ酸が増加すると云う報告のあることは注目される。
上原(1091)は、即浸冷蔵種の孵化の悪くなる前の、冷蔵期間25日頃から、卵内に遊離のプロリンが増加し、40-45日ではやや減少すること、ロイシン、フェニールアラニンも同様な動きをすることを報告し、高橋(914)もプロリン、グルタミン、グルタミン酸、ヒスチジンなどが冷蔵期間の長くなるのに連れて増加するのを認めた。高橋(915)は、また、冷蔵期間が長くなって障害の出初めた卵の磨砕液に熱水を加え、その遠心上清に1%のHgSO4またはHgCl2を滴下すると白色沈澱が生ずるが、 これは増加したプロリンその他のアミノ酸、あるいはそれに近い低分子の物質が沈澱するのであろうと考えた。
佐々木ら(809)は、蚕卵の遊離アミノ酸の研究中に、上記の現象に注目して実験を行なった。プロリン、ヒスチジンは、産卵直後(珍らしい取扱いである)から9日間冷蔵し、出庫当日浸酸した卵においても(浸酸直後)、冷蔵前に比べて著しく多かったが、プロリンはその後、反転期まで増加を続け、発蟻前に幾分減少し、ヒスチジンは発蟻まで増加を続けた。しかし、プロリンは浸酸および冷蔵を施さない卵*の反転期にも著しく多かった(催青著手前の測定は行なっていない)。プロリンソ、ヒスチジンの少ない産卵直後の卵の除蛋白濾液を加水分解すると、これらのアミノ酸が増加するから、産卵直後の卵にはプロリン、ヒスチジンなどを構成要素とする低分子のペプタイドまたはアミノ酸誘導体が存在し、卵内容の異常な組織分解によって、それらのアミノ酸の増加を来たすのであろうと云う。
(註:*1955年夏蚕用の秋花×銀嶺であると云うが、春採ならば浸酸しなければ反転期まで進まない筈であるし、
もし越年種ならば冬期の冷蔵を経ている筈であるから、この意味が不明である。)
化学の立場からではなく、蚕種の立場で問題点を生化学的に検討し、蚕種の取扱いとその結果とを結ぶ過程を明かにすることは今後心要な研究方向である。
催青中の卵を35℃に1日間接触させると2割程度の死卵が生じ、3日間の接触で殆ど全部が死卵になるから、蚕卵発育温度の最高限度は、実用的には35℃と考えられる(580)。
高温に対する抵抗力を胚の発育段階別にみると、小針らが(398)即浸種を40℃(湿度75%)に24時間接触させて孵化を調べた結果では、胚帯が卵表から離れて卵内に落込む頃(浸酸1時間後)、気管形成の始まった頃(浸酸当日を1日として数えた6日目)、頭部着色の頃(同7日目)および漿膜を飲み込む頃が弱く、中田(668)が不越年種を46℃-48℃(湿度20-35%)に20分および40分間接触させた結果では、分裂核分散、気門発生、反転(前期の第1期および第2期)および気管螺旋糸発生の各期が弱かった。越年種の催青中に45℃(湿度50-55%)に30分間接触させた木下(380)の成績では気門形成期、反転期および点青期が弱かった。反転の前後および点青期頃が共通的に弱い。
蚕種の抵抗力については、蚕種取扱いの実際問題として、以下の諸章において更に述べる。産卵直後の卵に対する高温の影響(Ⅳ5Aa)および休眠と保護温度との関係(Ⅴ1Ba)については既に述べた。
b 湿度
催青中の湿度が低いと孵化に悪形響がある。松村ら(514)の成績(第105表)によると、60%区は75%区および90%区に比べて明かに孵化歩合が劣っている。75%と90%との優劣は明かでなかった。7品種平均の催青日数は、25℃の場合に60%区11.33日、75%区10.98日、90%区11.00日で、75%および90%は60%に比べて短かかったが、75%と90%との差は明かでなかった。
| 蚕 品 種 | 催 青 湿 度 | ||
| 60% | 75% | 90% | |
| 日 1 号 支 4 号 欧 9 号 正 白 新 白 支4号×正白 新白×欧9号 |
78% 74 73 77 92 93 86 |
80% 78 89 83 95 92 88 |
83% 76 86 87 93 98 89 |
| 平 均 | 82 | 86 | 88 |
道家(88)によれば温度と湿度とは相関連して催青中の卵に影響する。総孵化歩合について云えば、95%以上の孵化歩合の得られる温湿度は15-28℃、50-95%の広範囲に亘るが、実用孵化歩合85%以上の得られる範囲は24-28℃、70-90%で、最適条件は25℃、85%であると云う。
孵化の際の湿度に関しては興味のある実験がある。室賀(613)は、最初の発蟻をみたときに卵面に桑葉を撒布すると、無撒布の対照区に比べて孵化が促進され、桑の乾葉を撒布しても同様の効果があるから、この場合の促進効果は湿度の影響ではなく、桑の香気の影響であるとした。これに対し、福田・橘田(118)は、さくら、いちょう、ぽぷら、のざわ菜などの葉を撒布しても桑と同様の効果がある;乾葉撒布では、室賀の結果とは違って効果が少ない;湿度を100%にした容器内においては、桑その他の生葉または乾葉を入れても殆ど効果がない;定湿にしたデシケーター中に1蛾の催青卵を分割して入れ、午後10時から翌朝10時までおいて孵化を調べると、第106表の結果が得られた、などのことから、上記の孵化促進効果は桑の香気によるものではなく、湿度の影響であろうと結論した。
| 蚕品種 | 湿 度 | |||
| 約17% | 50-60% | 70-80% | 100% | |
| 太 平 長 安 |
15% 18 |
16% 21 |
42% 46 |
71% 76 |
室賀・丸山(623)は、春採り越年性卵(欧7号および正白)を産卵後24時間目から湿度60%と80%との2区に分け、産卵後70日間は平均27.14℃(25℃以上30℃以下)、その後50日間は5℃、次いで0℃に25日間(冷蔵中の温度は両区同一とあるが、詳細の記載なし)保護した後、磨砕して生体含有水分率を比較した。生体含有水分率と云うのは卵殻を除いた部分の水分百分率であるが、5回の測定結果の平均は、正白80%区71.94±0.12%、60%区71.87±0.18%、差0.07±0.14%、欧7号80%区72.13±0.16%、60%区71.88±0.08%、差0.22±0.13%で、有意の差がなかった。蚕卵の生体重(対1,000粒)は10回平均が、正白80%区472.1±4.9mg、
60%区451.6±4.4mg、差20.5±1.2mg、 欧7号80%区634.6±4.9mg、
60%区618.8±5.1mg、差16.0±1.0mgで、何れも80%区の卵が重かった。(註:計算が合わない)。
産卵直後の多湿は蚕卵に対して有害で、特に温度の高い場合および低い場合にはその害が著しい(第107表)。
| 日 数 | 温度 | |||||
| 30℃ | 25℃ | 20℃ | ||||
| 湿度 | ||||||
| 75% | 98% | 75% | 98% | 75% | 98% | |
| 3日 5 10 20 |
97% 97 97 89 |
95% 94 87 56 |
94% 94 97 93 |
94% 91 76 41 |
95% 95 95 93 |
96% 95 91 58 |
山口・小林(1163)は、大シヤーレに硫酸、食塩水および水を入れて湿度0-100%の7区を作り、蚕種を入れた無蓋の小シヤーレをこの中に納めて越年種の冬期保護試験を行なった。供試したのは欧18号および改安の春採りで、10月23日から翌年4月10日までを上記の湿度区に分けて保護し、その間、毎月1回、卵の新鮮量、乾物量および孵化歩合を調べた。温度は自然温度である。
卵の新鮮量は何れの区においても、保護中に漸次減少し、減少の度合は低湿区ほど大きかった。乾物率はこれと反対に、低湿区では増加し、多湿区では減少の傾向を示した。
生物体にとっては、乾物量と水分量との一定の割合、即ち水分平衡を保つことが生理上望ましいものとするならば、乾物率の増減の最も少ないのが最もよい環境湿度と云うことになると想定したのであるが、このような湿度は、改安に対しては75%、欧18号に対しては50%であった。しかし、実際に孵化歩合の最もよかったのは90%区で、次ぎが75%および対照区で、最も悪かったのは、改安では0%区、欧18号では100%区であった(第108表)。
| 蚕品種 | 湿度 (%) |
孵化歩合((%) | |||||
| 11月 | 12月 | 1月 | 2月 | 3月 | 4月 | ||
| 改 安 | 0 10 30 50 75 90 100 対照 |
44 42 43 42 51 49 39 46 |
48 55 58 50 88 86 73 77 |
38 56 53 57 78 89 73 75 |
33 41 39 43 60 77 64 76 |
35 46 29 40 71 69 64 65 |
18 32 34 30 62 63 50 60 |
| 欧18号 | 0 10 30 50 75 90 100 対照 |
2 1 4 6 0 4 0 3 |
71 61 62 72 65 65 58 66 |
83 82 83 87 87 83 70 90 |
86 91 85 87 93 95 76 87 |
90 87 88 96 91 95 72 89 |
81 82 88 86 90 92 69 94 |
胚の発育は一般に多湿区が早く、また揃っていた(第109表)。
| 湿度 (%) |
蚕品種 | |||||
| 改安 | 欧18号 | |||||
| 調査時期 | ||||||
| 2月 | 3月 | 4月 | 2月 | 3月 | 4月 | |
| 0 10 30 50 75 90 100 対照 |
乙A-乙B 〃 〃 〃 〃 〃 〃 〃 |
乙A-乙B 〃 〃 〃 乙B 〃 〃 〃 |
乙B-丙A-丙B 〃 〃 〃 丙A-丙B 丙B 〃 〃 |
乙A-乙B 〃 〃 〃 〃 〃 〃 〃 |
乙A-乙B 〃 〃 〃 〃 乙B 〃 〃 |
乙A-乙B 〃 〃 乙B-丙A 丙A 〃 乙B-丙A 丙A |
試験全体を通じて、改安は区間の差が明瞭で、特に50%以下の乾燥に弱い傾向があり、欧18号は乾燥に強く、寧ろ100%のような多湿に弱い傾向を示した。この相違については、卵の大小、卵殻の厚さ、卵内容の相違なども考慮しなければならないとしている。
結論としては、冬期間(1-6℃)の蚕種保護の適湿は、催青の場合と同様に、75-90%であると云うのであるが、乾物率の増減を通して考えられる蚕種の代謝作用と保護環境との関係については論議が尽されていないように思われる。
なお、活性化の早さおよび全期間を通じての孵化歩合から考えて、改安は正常な状態の蚕種ではなかったように思われる。
催青温度と化性との関係についてはⅤ1Aa参照。
c 光線
渡辺(1148)によれば、点青期までの胚の発育は暗所におけるよりも明所において幾分速いが、その差は催青温度が20℃の場合に明かであって、25℃の場合には明瞭でない。点青期以後になるとこれとは反対に、明所におけるよりも暗所において胚の発育が進み、その差は低温よりも25℃以上の高温において明瞭である。更に発育が進んで孵化の段階になると、暗所のものは明所のものに比べて約9時間孵化がおくれる。これが、発育の進んだものの孵化を抑えて発育を揃え、一斉に発蟻させる暗催青法の基礎になっているのであるが、光に対する感受性は蚕の品種によって著しく相違している(421,429,954)。
恒明、恒暗および明暗交替(18時間毎)で催青した蚕種の孵化を比較すると、孵化は明暗交替区が最も早く、恒明区がこれに次ぎ、恒暗区が最もおくれるが、25℃恒明で催青した蚕種が若干孵化し初めたときから明暗の条件を種々に変えてみると、第110表のような成績が得られた。
| 試験区 | 光の条件 | 蚕品種 | |||||||||||
| S | N | ||||||||||||
| 経過時間 | |||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 | 5 | 6 | ||
| 1 2 3 4 5 6 7 |
150lux 恒明 暗・明(150lux)1時間交替 恒暗 5lux 恒明 5lux・30lux1時間交替 30lux 恒明 30lux・150lux1時間交替 |
151 0 0 12 15 218 179 |
162 138 0 31 243 151 159 |
31 3 0 27 23 48 20 |
38 146 0 17 48 50 61 |
36 2 0 13 27 30 27 |
35 89 0 32 63 41 64 |
94 69 66 99 79 103 101 |
54 41 59 41 38 19 46 |
33 29 12 46 32 30 39 |
23 36 8 27 15 17 25 |
15 14 6 12 13 15 14 |
10 24 8 17 12 13 14 |
これによってみると、光感受注の高い品種Sにおいては、恒暗にすると6時間の観察中、孵化が完全に抑えられた。5luxは恒暗と恒明との中間の作用をし、30luxでは150luxと殆ど同じ程度の効果があったが、150luxに比べれば明の効果が幾分弱いようである。Nは光感受性が非常に低い。
点青期以後を暗黒にしておき、若干孵化し初めたときから5、30、90、450luxの光を各1分間、5分間および450luxの光を60分間照射し、以後再び暗くして暗中での各区の孵化の遅速を比較しても(第111表)、NとSとの違いは明瞭である。
| 照射時間 | 蚕品種 | 調査項目 | 光の照度(lux) | 450lux 60分間 |
||||
| 0 | 5 | 30 | 90 | 450 | ||||
| 1分間 | N | 総孵化卵数(粒) 初め1時間の孵化歩合(%) 同上指数 |
65 66 100 |
67 67 102 |
58 81 122 |
69 65 98 |
49 57 86 |
56 83 127 |
| S | 総孵化卵数(粒) 初め1時間の孵化歩合(%) 同上指数 |
450 58 100 |
194 63 109 |
192 71 123 |
238 79 137 |
288 80 139 |
322 98 170 |
|
| 5分間 | N | 総孵化卵数(粒) 初め1時間の孵化歩合(%) 同上指数 |
271 60 100 |
202 70 117 |
160 61 103 |
247 65 109 |
154 64 108 |
148 75 126 |
| S | 総孵化卵数(粒) 初め1時間の孵化歩合(%) 同上指数 |
388 31 100 |
361 62 197 |
449 66 210 |
542 73 232 |
549 69 220 |
445 90 286 |
|
Sについてみれば、1分、5分ともに90luxと450luxとでは殆ど差がない。450lux・60分区の孵化の多いのをみると、照度だけではなく時間の影響することがわかる。孵化の誘発と云うことだけならば5lux・30秒でも効果は認められるが、実際問題としての孵化の促進には不十分である。しかし、発蟻抑制のための暗催青中にはこの程度の光でも抑制効果の妨げになる。本実験においては、調査のための蟻蚕の掃落しは写真用安全燈の下で手早く行なった。
以上のように、蚕卵の孵化は明暗によって著しく影響を受けるが、これとは別に、恒暗で催青してそのまま孵化させても、または恒明で催青したものを孵化前に恒暗にして暗中で孵化させても、一定の時間々隔で集中的に孵化の起こる現象がある。この孵化の週期(リズム)については多くの研究がある(985-994,1021-1027,1031)。
漿膜細胞の色素粒は低温によって集合し、温度の上昇によって分散するが(Ⅵ3Da)、光線の照射はこの分散を促進し、3・ヒドロキシキヌレニンからの漿膜色素の形成をも促進するもののようである。また、光は、点催青期の卵においては蟻蚕の斑紋色素の形成を促進し、発生初期には斑紋色素を減少させる傾向もあると云う(635)。
d その他の条件
点青期の蚕種を種々な気圧のもとにおくと、+100mm、-50mm、-100mmのような比較的小さな変化の場合には、常圧区よりも死卵歩合の減少する傾向が認められるが、その影響は処理時間6、24、48時の間では差がなかった(696)。また、点青初期の蚕卵を常圧より30mm低い気圧の下に20時間および40時間おくと常圧区に比べて、総孵化歩合に変りはないが、酸素消費量が増加し、孵化の促進がみられたと云う報告もある(1199)。
しかし、点青以前の蚕卵を-20mmで3日、5日、8日などの長期間処理すると催青日数が長くなり、孵化が劣るので、1蛾の卵を3分し、標高の違いによってこの程度の気圧差のある3カ所(東京、松本、小淵沢)に分けて催青試験を行なったが、孵化歩合に一定傾向の差が認められなかった(1198)。 但し、これらの試験においては、空気の組成などを調べてはいない。
長田(640)は0.5m/sec または1m/sec程度の気流があると催青日数が僅かながら短縮することを認めた(第112表)。しかし、孵化の際には妨げになるもののようで、卵殻に穴をあけたまま死んでいるものがやや多かった。
| 関係 湿度(%) |
気 流 |
催青4日17時間の胚 | 点青期に達した胚 | 催青日数 (日) |
孵化歩合 (%) |
|||||
| 調査卵 数(粒) |
反転前 のもの (%) |
反転中 のもの (%) |
反転後 のもの (%) |
調査卵 数(粒) |
催青8日 12時間に おいて(%) |
催青9日 12時間に おいて(%) |
||||
| 25 | 有 無 |
109 101 |
39 62 |
25 17 |
36 21 |
564 528 |
26 1 |
84 63 |
12.0 12.5 |
66 68 |
| 75 | 有 無 |
100 100 |
37 44 |
15 29 |
48 27 |
627 581 |
35 24 |
88 87 |
11.2 11.7 |
80 81 |
| 100 | 有 無 |
111 101 |
32 47 |
14 18 |
53 35 |
522 582 |
27 7 |
91 90 |
11.4 11.7 |
80 83 |