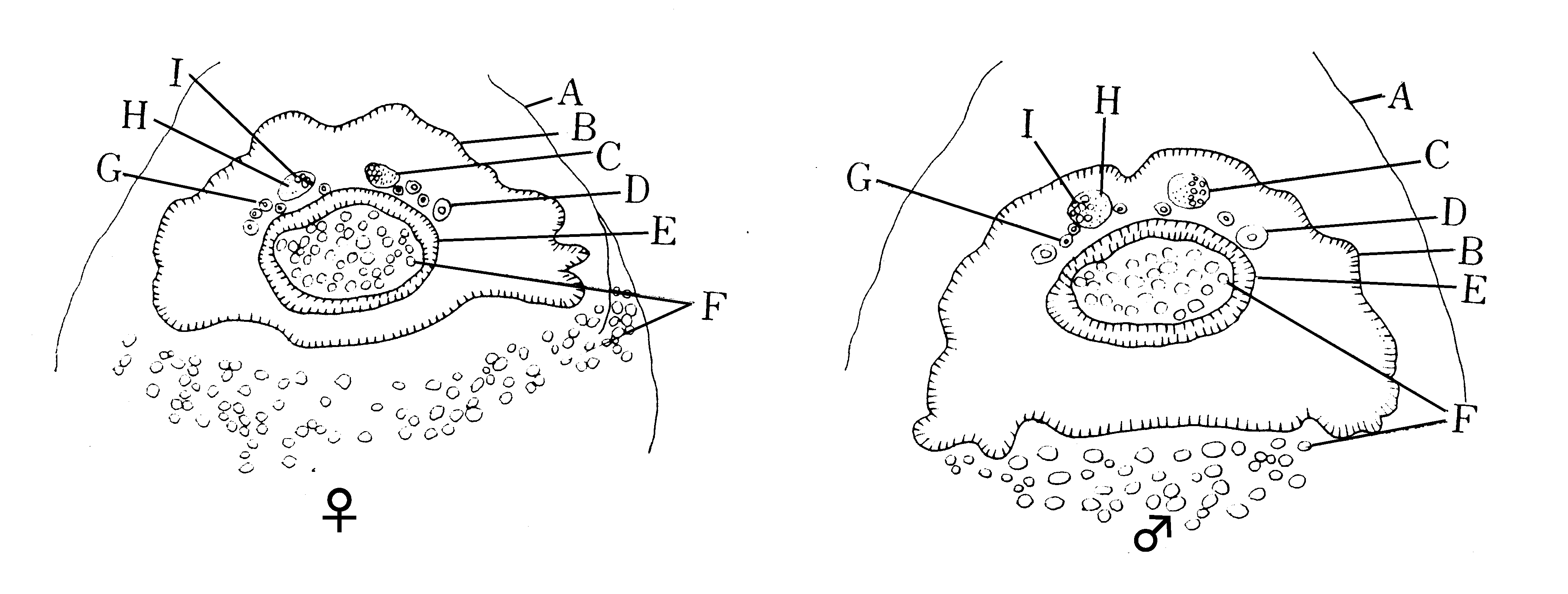
第1図 完成胚の断面図(石渡(261)模写)
A:漿膜、B:外皮、C:生殖器、D:絹糸腺、E:胃壁
F:卵黄、G:マルピーギ管、H:小細胞部、I:大細胞部
蚕 種 総 論
著者:高見丈夫
発行:全国蚕種協会
(昭和44年5月31日)
発刊のことば
蚕種取扱いの適否が蚕作に大きな影響を及ぼすことは今更言うまでもない。われわれ蚕種製造業者は蚕種の製造過程すなわち、原蚕飼育、種薗保護、採種、浸酸、冷蔵保護、催青等のすべてに亘り、長期間細心の注意を払い、その蚕種が養蚕家の手許に健全な状態で渡り、りっぱな繭が出来上るまで片時も気をゆるすことは出来ない。
従ってわれわれは蚕種の製造技術の全般にわたって、つねに研鑽を怠らず新しい技術の吸収に心を砕いている。近時おびただしい研究成果が発表せられ、これらを基とした新しい参考書の上梓を渇望していたのであるが、茲にに高見博士の力作「蚕種総論」の発刊を見てわれわれは百万の味方を得たような心強さを覚ゆる次第である。
高見博士は蚕種に関する学理と応用の両面に亘る真摯な学究として知られていたが、先般農林省蚕糸試験場を退職され小閑を得られていることを承り、同博士にわれわれの希求している参考書を執筆していただくよう前蚕糸試験場長大村清之肋博士を通じ、再三に亘り懇願したるところ、漸く御承引を得て今回茲に「蚕種総論」刊行の運びに至ったことは私の最もよろこびとするところであります。
ここに、執筆者高見丈夫博士の御努力に対し心から敬意を表すると共に、同氏の蚕種に関する研究成果があますところなく本誌に織込まれておるので、研究者、製造実務にたずさわる技術者はもちろん広く関係者の皆さんの御期待に添うものであることを確信して発刊のことばと致します。
昭和44年6月
全国蚕種協会会長 矢沢 泰明
序
わが国の蚕種に関する研究報告は尨犬な数にのぼっている。これだけの資料があれば、本来ならば、蚕種の技術はもはや名人芸をはなれて、電子計算機にたよれる可能性もあって然るべきであろう。しかしそれらの試験成績は、正しい吟味を尽されることなく放置されているので、これらの資料をそのまま電子計算機に覚えこませたら間違った答の出る心配の方が多い。
私は戦後蚕糸試験場に入り、はじめて蚕種の研究にたずさわったのであるが、水野、渡辺、梅谷、高梨等の蚕種学の泰斗の本拠であったここにおいてさえ、これらの方々の考え方の基本が伝わっているとは限らず、しばしば困惑したことがある。研究の内容が真には理解されないままに方法だけが伝わっている場合が多いので、例外的な条件下では正しい蚕種の取扱いができず、思わぬ失敗をすることがあったのである。つまり研究によって生まれた原理が理解されてもいないし、当然伝えられてもいないところに問題があるわけである。
研究業績の数がいかに多くとしも、その中に内在する原則的なものを掴まないと技術の進展は望み難い。本書は今までの多くの研究業績をできるだけ吟味して蚕種に関する現在の技術指針をたて、あわせて将来の研究推進の方向づけをしようとしたものである。もし本書が読者に対して、蚕種に関する多くの研究業績の内容を紹介するだけにとどまらず、蚕種に関する諸技術について解明すべき問題点を見出されたり、新らしい考えを持たれることに役立てば著者の喜びはこれにすぎるものはない。
本書に引用した表の数字は、解釈上支障がないと思われる場合は、小数点以下を省略した。また表の見出しを変更したり、表の一部だけを借用したりした場合や原著者とは違う論議の材料に用いた場合もある。従って、表の数字や解釈について疑義の出た場合の責任はすべて著者にあって、原著者にはないことを断っておく。なお古い品種名についている“国蚕”は記載を簡単にするために省略した。外国の文献は実際上、その必要はないと考えたから使用しなかった。
本書は大村博士のおすすめと、全国蚕種協会の御支援によって稿を起こすことになったものである。心から感謝を申上る。あわせて、種々御力添えをいただいた蚕糸試験場の方々および業績を勝手に使用させていただいた多くの研究者に御礼と御詫を申上るものである。
1969年4月
高見 丈夫
目次
| Ⅰ 家蚕の生殖器官 | ||
| 1 生殖腺の発生 2 雌の生殖器官 A 卵巣 a 幼虫 b 蛹 i 卵巣の発育および造卵数 ii 卵黄および欄間区の形成 c 蛾 |
B その他の器官 a 内部生殖器官 b 外部生殖器官 C 雌生殖器官の異常 a 正常型の個体変異 b 遺伝的な異常 c 偶発的な異常 |
3 雄の生殖器官 A 睾丸 a 睾丸の発育および精子の形成 b 無核精子及び精子の長さ B その他の器官 a 内部生殖器官 b 外部生殖器官 C 雄生殖器官の異常 |
| Ⅱ 原蚕飼育と造卵および蚕卵 | ||
| 1 体内環境と造卵および蚕卵 A 蛹重 B 卵の造成と絹物質の造成との関係 |
2 飼育環境と造卵および産卵 A 飼料 a 葉質 b 給桑量 c 葉質および給桑量と次代蚕 |
B 飼育環境 a 温湿度 b 光線その他 C 条桑育および屋外育 |
| Ⅲ 上蔟・種繭保護と造卵および産卵 | ||
| 1 上蔟・吐糸 2 卵母細胞および蛹の発育階程 3 蛹の保護 A 蛹の保護 a 保護環境と蛹期間 b 化蛹までの蔟中保護温度と蛹期間 c 発育段階別保護温度と蛹の発育 |
B 保護温度と造卵および蚕卵 a 造卵数、産卵数 b 保護温度と卵の代償 c 保護温度と卵の越年性 d 保護温度と孵化の良否 i 越年種 ii 人工孵化種 |
4 発蛾調節のための蛹の保護 A 温度管理 B 光線管理 5 蔟中および蛹の保護環境と各種の障害 A 温湿度による障害 B 炭酸ガスによる障害 |
| Ⅳ 採種 | ||
| 1 雌雄分離 A 性徴による雌雄鑑別 B 限性遺伝の利用による雌雄鑑別 C 繭重による雌雄鑑別 D 蛹の体幅による雌雄鑑別 E その他の方法 a 物理的方法 b 孵化の早晩と雌雄 2 発蛾 A 発蛾の行動 a 発蛾および脱繭 b 脱繭不能蛾 |
B 発蛾の早晩 a 発蛾の早晩と産卵 b 発蛾の早晩と化性 3 交尾 A 交尾時間 a 交尾時間と受精 b 交尾時間と産卵 B 雄蛾の使用回数 C 交尾中の管理 a 温度 b 光線 |
4 蛾の抑制 A 雌蛾の抑制 B 雄蛾の抑制 5 産卵 A 産卵中の管理 a 温度 b 光線 c 産卵不良蛾 性状 d 産卵順序および産卵状態と蚕種の B 産み着け場所 a 産み着け面の角度 b 産み着け面の性質 c バラ種台紙用糊料 |
| Ⅴ 化性 | ||
| 1 越年性の管理 A 化性の変化 a 温湿度および光線による変化 b 遺伝性 |
c その他の原因による化性変化 B 休眠 a 休眠の完成 b 休眠の解消(活性化) |
2 休眠の生理 A 抑制質 B 休眠に伴なう卵内の変化 C 休眠ホルモン(休眠要因) |
| Ⅵ 蚕卵の形態、発生および生理 | ||
| 1 受精前の蚕卵の形態 2 発生 A 受精 B 胚の発育階程 a 前休眠期 b 休眠期 c 越冬期 |
d 臨界期 e 器官形成期 f 完成期 3 生理 A 水分 B 呼吸 a 不越年性卵 b 越年性卵 |
C 発育 a 胚の成長 b 卵重の変化 D 環境 a 温度 b 湿度 c 光線 d その他の条件 |
| Ⅶ 越年蚕種の保護・取扱い | ||
| 1 春採り蚕種 A 蚕卵から休眠の完成まで B 休眠完成から初冬期まで C 人工温度について D 越冬期 a 単式冷蔵法 |
b 中間手入れ式冷蔵法 c 複式冷蔵法 2 初、晩秋および初冬採り蚕種 A 初、晩秋採り蚕種 B 初冬採り蚕種 |
3 蚕卵の簡易解剖法および孵化調査の注 意 A 蚕卵の簡易解剖法 a 熱湯固定による方法 b 生体解剖法 B 孵化調査の注意 |
| Ⅷ 人工孵化 | ||
| 1 浸酸成績に影響する一般的な条件 A 蚕卵の条件 B 塩酸の条件 a 夾雑物 b 濃度の調整 c 浸酸による塩酸濃度の変化 C 塩酸液の濃度と浸漬時間との関係 D 塩酸液の温度と浸漬時間との関係 2 即時浸漬 A 加温即時浸漬 a 浸酸 |
b 浸酸前冷蔵 c 脱酸、乾燥 d 比重1.10の塩酸液による加温浸酸 B 常温即時浸酸 C 不越年種および即時浸酸種の冷蔵 a 不越年種 b 加温即時浸酸種 c 常温即時浸酸種 3 冷蔵浸酸 A 冷蔵浸酸の基本問題 B 冷蔵浸酸の施行標準 |
a 冷蔵期間40日以上の場合 b 冷蔵期間40日未満の場合 C 冷蔵浸酸種の再冷蔵 D 浸酸奏功の判定 4 浸酸および浸酸後の冷蔵による蚕種の 障害 5 随時孵化および冬掃き蚕種の浸酸 A 随時浸酸 B 冬掃き蚕種の浸酸 6 人工越冬 |
| Ⅸ 催青 | ||
| 1 催青の準備 A 予備催青 B 催青室および催青用具の準備、消毒 C 蚕種および蟻蚕の消毒 |
2 催青条件 A 温度 B 光線 C その他の条件 催青 D 二化性種を不越年化させるための E 催青条件と眠性の変化 |
3 催青中の蚕種の冷蔵および蟻蚕の抑制 A 催青中の蚕種の冷蔵 B 蟻蚕の抑制 C 蟻蚕の2夜包み、3夜包み |
| Ⅹ 蚕種の調製および輸送 | ||
| 1 洗落し A 洗落しの時期および水温 B 比重選 C 蚕種の膠着防止 |
2 卵量と蟻量 3 蚕種の輸送 |
|
| XI 蚕種の事故 | ||
| 1 事故蚕種の形態と原因 A 無着色卵 B 着色死卵 a 催青着手前の越年種にみられるもの b 越年種の催青を始めてから発現する もの c 人工孵化種にみられるもの C 淡色卵 |
D 白ハゼ卵 E 卵液の滲出 F 卵黄膜の剥離 G 点催青死卵 H 不孵化卵 I 卵殻喰破りの異常 J 再出卵 K 孵化不斉 |
L 蟻蚕の畸形 M 死蟻 2 病毒の経卵伝達 A ウイルス病の経卵伝達 B 経卵伝達の証明 C 蚕種の実際問題 D 蚕種の立場からの対策 |
Ⅰ 家蚕の生殖器官
1 生殖腺の発生
蚕の卵や精子の起原である生殖細胞は内胚葉から分化すると考えられたこともあるが、外山(1074)は、内胚葉ではなく中胚葉起原であるとし、中胚葉が2層になったときその内層(1902年の論文(1075)には外層に当るSomatic mesoblast から分化すると書いている)から分化することを記載した。これは胚に付属肢突起の形成され初めた時期に当る。しかしその後、川口・宮の報告(331)に続き、宮が多くの研究結果を発表し、生殖細胞の分化、出現はこれよりも遙に早いことを明にした。その結果を要約(561,562)すると次ぎの通りである。
不越年性卵においては、後(Ⅵ)に述べるような発生経過を経て、産卵後16時間ぐらいで胚帯が形成される。このとき、外形は他の細胞と余り違わないが、核が幾分大きく、はっきりした大形の仁を有する細胞群が胚帯の一定の部分から部内に向って陥入する。これが初めて観察される生殖細胞の姿である。生殖細胞としての分化はこの陥入より以前に起っているものと考えられるが、形の上ではこのときまで区別ができない。
陥入した生殖細胞は胚の内面に沿って前後に分散し、胚の頭部に沿うあたりまで分布するものもあるが、産卵後24時間目頃になると腹部に沿って分布しているものは再び胚帯の中に取込まれ、頭胸部に沿って分布するものは取込まれずに胚外に残る。これは所謂“だるま形”胚の時期である。次いで、胸部から中胚葉の陥入が始まり前後に進行するが、これに伴なって、腹部では、生殖細胞を取込んだ部分が生殖細胞を含んだまま中胚葉として陥入する。産卵後36時間ぐらいで中胚葉が体節状に分れ、39時間で付属肢原基が生じ、口陥が陥入し、神経溝によって各体節の中胚葉が左右に2分されるが、これに連れて、含まれている生殖細胞群も2分される。
この頃から、それまで1層であった中胚葉組織が側端から2層になり初め、42時間で肛陥が始まり、顎節および胸節に付属肢が形成され、中胚葉の2層化も進行する。この2層の中胚葉の外層が体壁中胚葉、内層が内臓中胚葉であるが、生殖細胞は体壁中胚葉に包まれている。外山が初めに(1075)観察したのはこの頃の生殖細胞の状態ではないかと思われる。その後45時間目頃までの間に、中腸原基が現われ、肛陥の盲端に近くマルピーギ管が出現し、神経母細胞が分裂して神経球を作り、腹節の付属肢が形成され、気門も生じるが、生殖細胞には目立った変化がない。
最初に陥入する生殖細胞の数は蚕の品種によっても異なるが、多くても30個内外で(第1表)、出現後暫らくの間は分裂することなく、生殖腺形成の始まる産卵後70時間目頃まではその数が増加しない。従って、越年性卵においては、生殖腺形成の始まるのが休眠終了後であるため、産卵後7日たっても生殖細胞の数に変化がない(第1表A)。
| 蚕品種 | 産卵後 の時間 |
調査した 胚の数 |
生殖細胞数 | 生殖細胞に 分裂の認め られた胚の数 |
||
| 最少 | 最多 | 平均 | ||||
| p22 | 21時間 24 36 39 42 |
4 8 10 9 10 |
12 13 9 13 9 |
17 18 20 22 26 |
15 16 15 14 16 |
0 0 0 0 0 |
| br2 | 36 48 |
20 15 |
21 19 |
37 36 |
31 29 |
0 0 |
| SSY | 48 | 17 | 11 | 32 | 24 | 0 |
| g40 | 36 48 60 72 84 |
20 20 11 11 11 |
10 11 12 15 14 |
18 25 20 30 26 |
15 16 16 23 18 |
0 0 0 1 3 |
| A | 16 7日 |
8 8 |
9 8 |
14 18 |
12 12 |
0 0 |
昆虫の種類によっては、卵の特定の部分に特別の周辺細胞質があり、ここに入った分裂核だけが生殖細胞に分化するから(484)、そのようなものでは、生殖細胞が卵のどの部分で分化し、何個の原生殖細胞に由来しているかもはっきりわかるが、蚕卵の周辺細胞質には生殖細胞分化の場所として識別できる部分がなく、また胚帯から陥入してくるまでは生殖細胞を他の細胞から識別できないため、正常発生の観察によっては、上に述べた以上に生殖細胞の分化を追究することができない。
そこで宮は卵の種々な部分を部分的に焼き殺し、残りの部分から発生する部分胚について生殖細胞の発生を研究した結果、卵の中で生殖細胞の分化する位置は分裂核が周辺細胞質に到達する以前に既に決定していることを明かにした。この生殖細胞の分化予定域は、卵の長軸(前極と後極とを連ねる線)上、後極から30%内外の位置において、腹面正中線を挾んで左右何れも卵周の2.5%の範囲内にあり、この部分の周辺細胞質に入った分裂核だけが生殖細胞に分化するのであって、この分化予定域を焼き殺すと、胚は形成されても生殖細胞は分化しないことがわかった。発生が進んで胚盤期になると、予定域の範囲は幾分縮まるようである。
初めに陥入する生殖細胞の数は胚盤の完成までにきまるものと考えられる。川口(330)は、遠心力を作用させて作ったモザイク蚕の翌代を調べ、親の皮膚のモザイク状態と子供の表型との関係から考えて、原生殖細胞の数は最少限20個前後であろうと推定したが、その数が宮の観察した最初に陥入する生殖細胞の数とほぼ一致することは興味がある。
陥入後、胚の内面に沿って分布した生殖細胞のうち、腹部に沿って分布するものだけが胚帯に取込まれることを先きに述べた。その後、中胚葉と共に再陥入するが、その分布範囲は胚の長さが増大するとそのまま引きのばされるだけで、胚の各部における分布割合には変化がない。
産卵後60時間目頃から体腔嚢が破れ初め、やがてその腹壁が内方に伸び、その一部が生殖細胞群を取囲んで、胸腹部第6-9の各節毎に生殖隆起を作る。これが生殖腺形成の第一歩で、これと共に生殖細胞の分裂、増殖が始まる(第1表)。これは産卵後ほぼ70時間目頃であるが、その後、胚の発育に伴ない各節の生殖隆起は伸長し、前後互に連絡して体の腹面両側に索状の組織を作り、次ぎにこれが次第に第8節に向って前後から収縮し、78時間目の反転前期(Ⅵ2Be)には第7節の後端から第8節の範囲に縮まってくる。84時間で、生殖細胞とそれを包む上皮および後に紐体(第2、10図)を形成する細胞群が識別されるようになり、幼虫における生殖腺の形態がほぼととのうが、これが第8節背面に移動を終えるのは反転完了のときで、このときにはヘロルド腺も発生し(668)、紐体が第11乃至12節の腹面に連なる(1074)。
胚の生殖腺はその中に含まれる生殖細胞数が多いほど大きいが、含まれる生殖細胞数は種々な条件によって影響される。最初に陥入する生殖細胞の数が品種によって相違するのを(第1表)、宮は生殖細胞分化予定域の大きさの相違によるものと考えている。しかし、最初の生殖細胞数は同じでも、陥入後、腹部に分布したものだけが胚帯に取込まれ、更に第6-9節のものだけが生殖隆起に包まれて生殖腺形成に参加できるのであるから、かなりの数の生殖細胞が生殖腺形成から除外されることになる。正常の場合には、生殖細胞は第6-8節に最も多く分布するが、後方は第10節、前方は顎節の付近にまで分布するものがあり、宮は、全生殖細胞数の14%にも達する数が第6-9節以外に分布していた例をみている。この生殖細胞の分布はチャンスによって機械的にきまるものと考えられ、丁度陥入の起こる頃に卵の後極を焼くと前方(胸部)への分布が多くなると云う。更に、第6-9節に分布する生殖細胞も全部が生殖隆起に包含されるとは限らず、第6-9節に分布しながら、全数の36%もの生殖細胞が生殖隆起に含まれずに終った場合も観察されており、その原因の一つとしては、生殖隆起の形成に関与する中胚葉細胞の数が不十分で、総べての生殖細胞を包含し得ないことが考えられている。
第1図 完成胚の断面図(石渡(261)模写)
A:漿膜、B:外皮、C:生殖器、D:絹糸腺、E:胃壁
F:卵黄、G:マルピーギ管、H:小細胞部、I:大細胞部
以上述べてきた生殖腺は未だ卵巣か睾丸かの区別のできない時期のものであるが、石渡(260,261262)によれば、完成した胚においては、胚の横断切片標本を作って生殖腺の中央部を通る断面をみると(第1図)、左右の生殖腺の相対した側に小細胞(被膜細胞)、反対側に大細胞(生殖細胞)の集合しているものは睾丸、相対する側に大細胞、反計側に小細胞の集まっているのは卵巣であり、またヘロルド腺(第10図)のあるものは雄であるから、胚においても雌雄が識別できると云う。小細胞の集合している部分は紐体の付着点であるから、幼虫における紐体付着点の位置(第2、10図)から考え、順次にさかのぼって観察すればこのような結論になる訳であるが、生殖腺の形、大きさ、位置、紐体の付着点などは幼虫においてもかなり変異のあることからみて、一般的には上のように云えるとしても、個々の場合にはこの標準で正確な判別ができるか否か疑問がある。その上、固定切片にして調べるのであるから、卵を孵化させてその判別の当否を直接に確かめることもできなかった。現在は限性黒卵品種があるので、これを用い、予め雌雄を分けた卵を切片にして調べれば、上記の方法でどの程度に雌雄鑑別ができるかを確かめることができる筈である。
町田(490)は、孵化当初の幼虫においても、1)卵巣は睾丸より小さい; 2)紐体の付着点が睾丸では左右相対する側にあり、卵巣では反対側にある; 3)睾丸小胞の中央部には生殖細胞が殆ど存在しない; 4)卵巣小胞の先端には尖端細胞があるが、睾丸小胞にはこれが認められないなどの一般的標準では、卵巣と睾丸とを個々の場合に正確に識別しかねる場合の多いことを述べている。
なお町田は、卵巣においては尖端細胞が孵化当初既に認められるが、睾丸では孵化翌日になって初めてこれが現われ、卵巣におけるよりも顕著であると云う。しかし、宮(562)は尖端細胞と考えられる細胞を孵化直前の生殖腺で記載し、しかもこの生殖腺は紐体の付着点から考えて睾丸であろうとしているから、尖端細胞のあるなしも正確な雌雄鑑別の基準には用いられない。生殖細胞の数は孵化を境にして急激に増加する(第2表)。
| 時 期 | 調査した 生殖細胞 |
1生殖腺当りの生殖細胞数 | ||
| 最少 | 最多 | 平均 | ||
| 孵化2日前 1日前 当日 1日後 |
17 28 11 15 |
29 37 82 102 |
57 79 162 236 |
47 59 119 171 |
2 雌の生殖器官
A 卵巣
a 幼虫
幼虫の卵巣は第8節の背面、星状絞のある部分の皮下に、背脈管を挾んで左右に1対ある。孵化当時は睾丸との識別が困難であるが、日のたつに連れて、前節で述べた種々な特徴によって識別できるようになり、3令起蚕においては、卵巣はほぼ3角形を呈し、3角形の底辺で左右のものが相対している(第2図)。これに対し、睾丸は腎臓形で丸味があり、その凹面で相対し(第10図)、紐体の付着点も相違している。

第2図 幼虫卵巣の位置模式
a:卵巣、b:紐体、c:紐体末端、d:石渡前腺の位置
(背面からみたもので、田中(1035)によって描いた)
孵化当初の幼虫の卵巣には内部の区画が明瞭でないがやがて隔壁が発違して、普通4個の小胞状の区画に分れ、日を経るに連れてこれが伸長して小管状となり、更に伸長を続けそのままでは共同被膜内に納まらず、3令2日目頃から屈曲し初める。
蚕令に伴なう卵巣の大きさおよび卵巣小管(卵管)の長さの増大は第3図によっても明かであるが、町田(491)が小石丸品種について測定した結果を曲線で表わすと第4図のようになる。この場合、1令期間は切片標本による測定であり、2令以後は生体のまま解剖して測ったのであるから、同一に扱うことのできない測定値の接ぎ合わせであるが、便宜上1本の曲線で示した。日々の変動がかなり大きいが、これは測定数が少ないので重要視することはできない。全体の傾向として、卵巣の長さおよび幅の成長は、初めは徐々に、後次第に速く、5令期には更に速くなる。これに対し、卵管(卵管柄をも含めて測定した)は5令の終りまでは徐々に伸び、上蔟後急激な伸長を示し、化蛹2日目に共同被膜が破れて腹腔内に出た。その後も卵管の伸長はますます著しく、遂には腹腔内に充満する。

第3図 各令卵巣の大きさの比較(田中(1035)模写)
A:3令起蚕、B:4令起蚕、C:5令起蚕
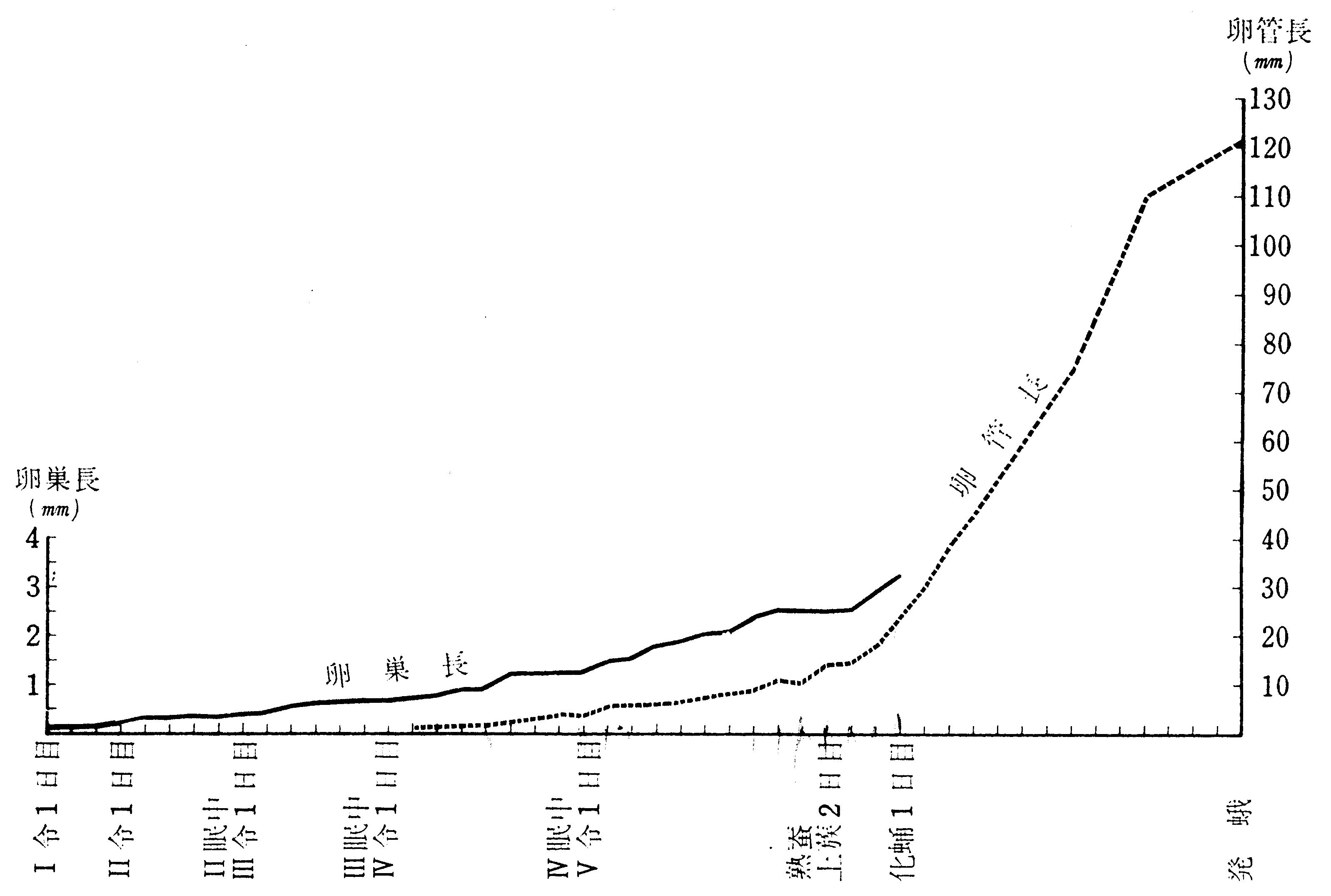
第4図 卵巣の成長曲線
(町田(491)の数字によって描いた)
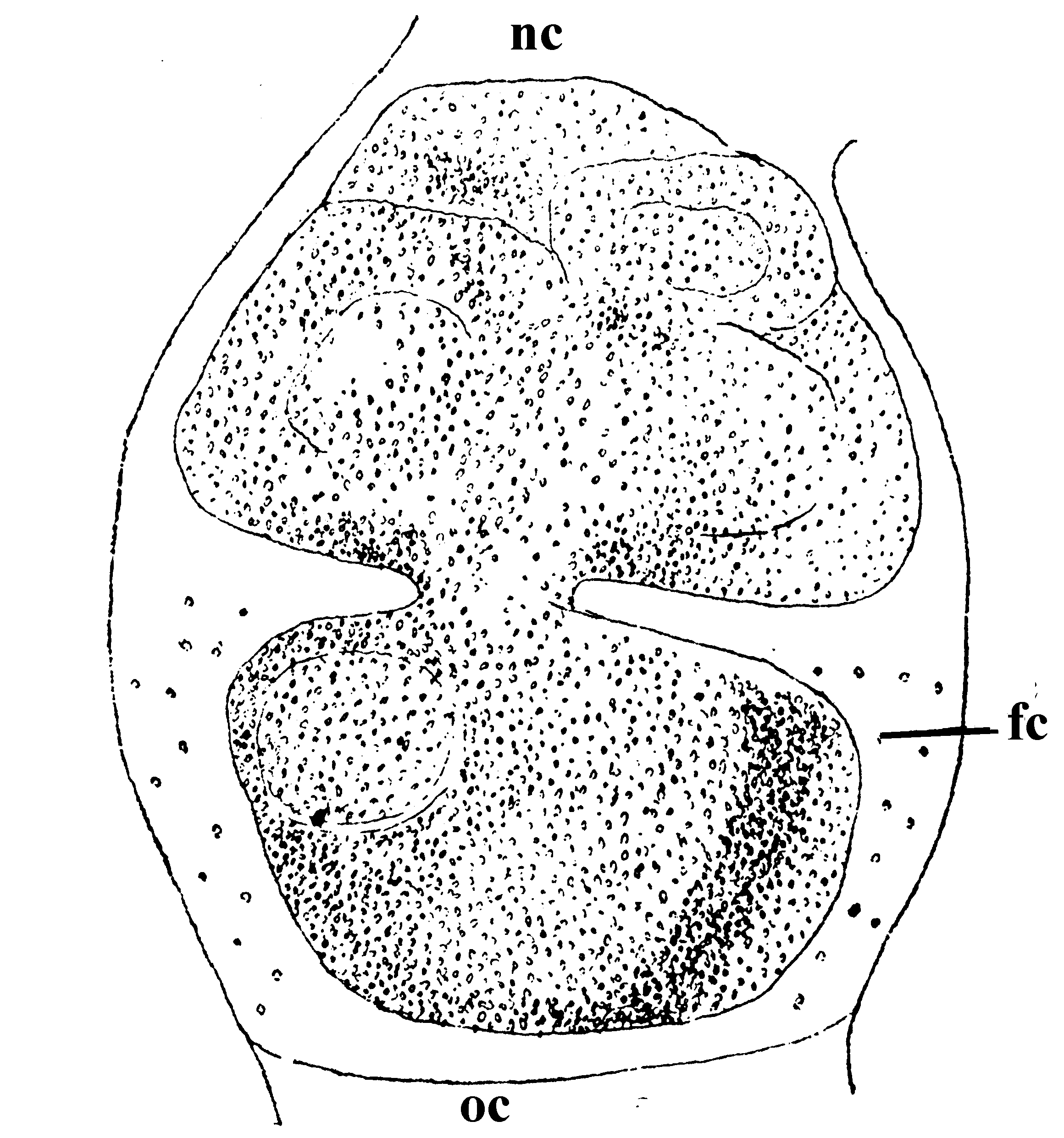
卵巣の外形がこのような変化をする間に、その内容にも重要な変化が起こる。孵化当時の卵巣小胞内には生殖細胞とそれに混って若干の包卵細胞があるだけである。生殖細胞(卵原細胞)は卵管の先端部で分裂、増殖を続けるが、順次に分裂を止め、次第に卵管内を下降して、卵母細胞と栄養細胞とに分化する。この場合、分裂を止めるものは、最後の分裂の3回前から、分裂した細胞同志が一団となって離れず、結局、8個ずつが1集団となり、そのうちの1個が卵母細胞、残りの7個が栄養細胞に分化するのである。分化の認められるのは4令2日目頃からで、栄養細胞に分化したものは生殖細胞としての機能を失い、卵母細胞に栄養を供結する役目を特つことになる。次いで、これらの細胞集団内で、卵母細胞と栄養細胞との位置的関係がきまり、4令中期には、卵母細胞が下部にきて、その上部に7個の栄養細胞の付着した一定の配列のものが現われてくる(第5図)。

第5図 配列のきまった卵母細胞(町田(500))
nc:栄養細胞、oc:卵母細胞
これに包卵細胞が加わって(第6図)、卵管内に、基部から先端に向い1列に珠数玉状にならぶのであるが、珠数玉の一つ一つに当る細胞集団は、上部に、栄養細胞の納まった栄養室、下部に、卵母細胞の納まった卵室を形成する。従って、卵管全体としてみれば、栄養室と卵室とが交互にならんでいるので、交互栄養型(多栄養型)卵管と呼ばれる。5令の初期には卵管の基部にこの交互栄養型の特徴が現われてくる。
第6図 卵室、栄養室の形成初期(町田(500))
fc:包卵細胞、nc:栄養細胞、oc:卵母細胞
卵母細胞と栄養細胞との間にこのような配列がきまっても卵の完成にはなお遠いが、卵は卵管の下方から順次上方へ成熟してゆき、産卵もその順序で行なわれるのであるから、卵母細胞と栄養細胞との位置的関係のきまった細胞集団の数を基部から先端に向って数えると、将来完成*あるいは成熟する卵、または産み出される卵の数が何時頃に準備されるかを知ることができる。この点を調べたのが第3表である。
| *厳密な意味での完成卵は第二成熟分裂(Ⅵ2A)の終った成熟卵であるが、卵殻が完成し、産み出されるものと外観的に同様な形態に発育した卵を、包卵被膜から離脱する(Ⅰ2Ac)と否とに拘らず完成卵と呼ぶことが多い。普通に造卵数と云われているのはこの意味の完成卵数(産卵したものがあればそれを含めて)のことである。 |
| 5 令 | 上 蔟 | 化 蛹 | ||||||||||||||
| 3日 | 4日 | 5日 | 6日 | 7日 | 8日 | 9日 | 熟蚕 | 2日 | 3日 | 4日 | 1日 | 2日 | 3日 | 4日 | 5日 | |
| 1卵管当り | 17 | 23 | 40 | 42 | 49 | 46 | 48 | 66 | 68 | 74 | 81 | 87 | 118 | 131 | 128 | 131 |
| 1個体当り | 139 | 187 | 318 | 332 | 395 | 371 | 381 | 527 | 545 | 592 | 651 | 698 | 944 | 1046 | 1022 | 1046 |
| 調査個体数 | 3 | 3 | 2 | 1 | 2 | 2 | 6 | 5 | 4 | 6 | 5 | 7 | 2 | 5 | 3 | 3 |
この調査の場合、化蛾したもの10頭について調べた造卵数は、最少546個、最多675個、平均618個であったから、この表によれば、それだけの数は既に化蛹の頃にきまっており、その後に増加した分は発育の過程で退化するか、または未発育に終ったことが明かである。化蛹5日日の卵数は1,049個となっているが、これは卵管鞘の外側からはっきり確認のでぎたものだけで、卵管のこれより上方には、分化はしても配列の未だはっきりしない卵母細胞が多数にあるから、これをも加えると、退化または未発育で終る卵母細胞の数は極めて多いことになる。これは造卵数の問題を考える場合に考慮しなければならない重要なことがらである。
永井(630)は、栄養細胞と卵母細胞との分化の明かになったものから卵殼の完成したものまでを含めた、数え得られる卵母細胞の数は、蛹期間の40-50%を経過したときに最大に達し、その後、化蛾までには大きな増減はなく、蛾になったときの完成卵数は全卵数の約80%であったと云っており、その完成卵歩合は第3表および第4表の場合に比べて著しく大きい。原因は明でないが、おそらく、分化した卵母細胞の基準が違うのであろう。
卵母細胞が分化すると卵黄形成が始まるが、これについては次項で述べる。
b 蛹
i 卵巣の発育および造卵数 蛹期における卵巣の発育は造卵数との関連上特に注目される。
蚕の産卵数は造卵数と造卵数のうちのどれだけが産み出されるかによってきまるから、造卵数の多いことが直ちに産卵数の多いことにはならないが、これが産卵数を決定する第一の要因であることには間違いない。卵母細胞は既に述べたような過程を径て分化し、発育するのであるが、蛹期間の中頃までの卵母細胞の数は、蚕の品種が異なっても飼育条件が相違しても大差がなく、また産卵数または造卵数と平行するものでもないらしい。例えば、長谷川(154)が日106号および支101号について調べた結果によれば(第4表)、日106号の化蛹5日目の卵母細胞数は第3表に示した小石丸の場合と同じであり、支101号はこれに比べて幾分多いが、ほぼ近似した数である。処が、総完成卵数は、小石丸が平均546個であったのに対して、日106号は688(86×8)個、支101号は560(70×8)個で、卵母細胞数の多かったものほど完成卵数が多いと云う訳ではない。
| 蛹 令 | 卵母細胞数 | 卵管本部の完成卵数(a) | 卵管柄に下降した完成卵数(b) | 総完成卵数(a+b) | ||||
| 日106号 | 支101号 | 日106号 | 支101号 | 日106号 | 支101号 | 日106号 | 支101号 | |
| 化蛹当日 2日目 3日目 4日目 5日目 6日目 7日目 8日目 9日目 10日目 11日目 12日目 13日目 |
102 98 111 134 131 131 163 164 1681) 1612) - - - |
153 159 158 175 179 183 208 1813) 211 - - - - |
68 49 27 14* |
65 40 7* |
13 31 54 72* |
10 30 63* |
81 80 81 86* |
75 71 70* |
また、5令3日目(熟蚕の3日前)に緑蚕上蔟させた日106号についてみると、蛾の1卵管当り完成卵数は対照区83±1.8個に対し緑蚕区52±1.1個で明かに緑蚕区が少なかったが、化蛹7日目における卵母細胞数は対照区165±3.6個、緑蚕区144±3.6個で大差がなかった。即ちこのときの緑蚕区の卵母細胞数は対照区化蛾後の完成卵数の2倍近くもあったのであるが、その後退化して、完成卵になったものは上記のように少なかった。退化卵の現われ初めるのは対照区では化蛾の3日前(13日目に化蛾)であったが、緑蚕区では4日前(12日目化蛾)であった。蛹の中期頃までの卵母細胞数はまた取扱いを著しく違えた区間でも大差がなかった。
蛹期における卵巣の重量は(第6表)初めは徐々に増加するが、蛹期間のほぼ60%を経過した頃から急増し、末期には殆ど変化がなく、大きく分けて3段の成長をする(154,295,317,665)。初めは卵巣重の絶対値が小さく、蛹重に対する割合が小さいため、第5表の例では、蛹体重と卵巣重との相関係数を求めても、日106号の化蛹8日目、支101号の化蛹7日目までは負価を示すことが縷々あって一定しなかったが、日106号は9日目で+0.526±0.229、以後は+0.501~+0.825、支101号は8日目で+0.649±0.183、以後は+0.504~+0.720となり、卵巣重が蛹重の30%ぐらいに達すると正の相関を示すようになっている。
| 蛹 令 | 日106号 | 支101号 |
| 化蛹当日 2日目 3日目 4日目 5日目 6日目 7日目 8日目 9日目 10日目 11日目 12日目 13日目 |
0.3% 0.5 1.0 2.7 5.5 10.9 16.4 24.5 29.4 30.6 32.7 32.3 48.4(蛾) |
0.5% 0.7 1.3 2.8 8.0 17.8 27.8 32.7 32.5 32.2 52.1(蛾) |
清水(840,841)は初め、5令蚕児の第8節における左右の気門の大きさとそれぞれの側の卵巣または睾丸の大きさとの間に正の相関のあることを認め、もし異個体間にもこのような関係があれば、外観によって卵巣の大小がわかり、実用上役立つであろうと考えたが、幼虫期の気門の大きさと化蛾後のその側の卵巣の造卵数との間には相関がなかった。中曾根(665)は、5令起蚕で、体の片側の気門(第3気門以下全部)を閉鎖するとその側の卵巣の発育の劣ることをみている。
造部数との関係で興味のあるのは片側の卵巣を摘出した実験結果である。橋本(175)は、5令期に片側の卵巣を摘出すると、残された卵巣の完成卵数がふえ、卵の形も大きくなることを認めた。吉川(369)は、5令初期の蚕から取出した卵巣を1個、同じ発育程度の他の蚕に移植すると、移植卵巣も宿主の卵巣も共に発育が阻害されるが、移植卵巣の発育が悪いと宿主の卵巣はほぼ正常に発育し、移植卵巣がよく発育すると宿主卵巣の発育が妨げられ、結局、卵巣の数は1個多くなっても、その宿主体内で形成される完成卵の総数は宿主本来の完成卵数に近い数に止まると報告した。これらの結果は、蚕の体内で完成卵を作るのに必要な栄養物質の量が一定しているために、これを消費して発育する卵巣の数が多ければ、発育を許される1卵巣当りの卵数が減少することを示しているものと考えられる。
摘出した残りの卵巣の重量増加を第6表に示す。この場合、摘出区の蛹重は、手術の影響によって、化蛹当日には対照区に比べて約2.5g(対10頭)も軽く、従って卵巣重も軽かったが、化蛹5日目から摘出区の卵巣重が急に増加し、対照区よりも重くなった。余分な栄養物質の影響が、先きに述べた卵巣重の急増期になって初めて現われ、それまでの発育不良とは無関係な重量増加を示していることは、造卵数が蛹中期頃までの卵母細胞数と平行しないことと考え合わせて注目される。残存卵巣のカクラーゼ作用には変化がないと云う(663)。
| 蛹 令 | 摘 出 区 | 対 照 区 |
| 化蛹当日 2日目 3日目 4日目 5日目 6日目 7日目 8日目 9日目(蛾) |
0.04g 0.06 0.12 0.42 1.06 1.85 2.03 1.93 1.93 |
0.05g 0.09 0.21 0.71 1.28 1.62 1.71 1.89 1.90 |
長谷川(154)の実験によれば、5令3日目の日106号から片側卵巣を摘出すると、残った卵巣の卵管が対照区のものより長くなり、化蛹7日目の卵数は1卵管当り140±4.0個で対照(第4表)に比べて寧ろ少ないぐらいであったのに、化蛹13日目に化蛾したものの1卵管当り完成卵数は100±2.0個で対照よりも多く、退化卵の出現も1日おそかった(化蛾2日前)。これは、片側卵巣の摘出によって栄養物質の量に余裕が生じ、退化卵となるべきものが完成卵に発育したことを示すものであろう。
第Ⅵ連関群に属する異常生殖腺蚕遺伝子(EGd)を持つ蚕の造卵数もこの点に関して興味がある。この遺伝子はホモ致死であるが、ヘテロで卵箇数を多くする。市川(270,271)がこの蚕の造卵数を調べた結果によると、5令期の毎日の絹糸腺重も繭質も同蛾区の正常姉妹蚕と異ならないのに、1蛾当りの造卵数は626±8.98粒で、正常姉妹の589±4.13粒に比べて多かった(α=1%で有意)。しかし、100mgの卵数は異常生殖腺蚕184±0.37粒、正常姉妹165±0.47で前者が多かったから(α=1%で有意)、1蛾当りの造卵量にすれば、異常生殖腺蚕341mg、正常姉妹356mgで,異常生殖腺蚕の方が却って少ない計算になる。これならば、母体の他の器官の生産または消費に影響することなく十分に生産がまかなえる筈である。このことから、問題は卵の数ではなく、母体と生産卵との量の関係であることがわかる。市川は異常生殖腺蚕の系統に大卵遺伝子(Ge)を入れても、大卵で造卵数の多い系統を作ることのできなかった理由をこの関係によって解釈している。
これらの結果から考えると、飼育や取扱いによる造卵数増加の問題は、結局、
1)母体内諸器官への素材の配分率を変更することができるか否か
2)配分率が一定の場合には、
a 素材の絶対量を増加することができるか否か(例えば、母体を大きくする、あるいは必要な栄養素の量だけを増加する)
b 卵の数と量との関係を変更することができるか否か(例えば、上記異常生殖腺養のように)
に帰着するものと思われる。アラタ体を摘出すると造卵数が減る(1175)、 エクディゾンその他の昆虫ホルモンを与えると造卵数がふえた(402,403,404)と云うような成績も、ただ造卵数だけの調査では不十分で、上に述べたような関係を考慮して吟味する必要がある。
退化卵がどのような機構で生じるのかは明かでないが、それが初めて出現する位置は、卵管内で必らず完成卵に近く、相当に発育した卵が退化し、これより卵管の盲端に近い若い卵の退化はおくれると云うから(155)、未熟卵の発育が全体的に停止するのではなく、発育の旺盛な卵、あるいは発育のある臨界期に達した卵に起こる何らかの栄養物質の不足が原因であろうと考えられる。
退化卵においては、ニンヒドリン陽性物質、燐、グリコーゲンなどが減少または消失するが、化蛹初期にグリコーゲンや燐を注射しても退化卵は発現する(155)る。 赤尾(31,37)によれば、蚕体内諸組織に含まれていた亜鉛が化蛹時に体液中に出て卵巣に集積する。卵巣を摘出しておくと、集積する組織がないため、化蛾前の蛹の体液中に合まれる亜鉛の量が対照の4倍にも増加し、マンガンも亦卵巣摘出によって体液中に著しく増加すると云うが、長谷川(156)の実験によれば、亜鉛、マンガン、鉄、銅などの金属イオンを蛹に注射しても退化卵の発現を防止できなかった。1令から5令まで毎日1回、亜鉛またはマンガンを添食すると熟蚕の卵巣が大きくなり、産卵数および卵重が増加したと云う報告もあるが(969,970,978)、産卵数についてはなお検討する必要があるように思われる。
ある物質が卵母細胞の発育と時期を同じくして蛹体内で著しい消長を示す例は、亜鉛やマンガンに限らず、ビタミンB2にもみられる(89)。雌の体液中のビタミンB2含量は蛹期の初めに急増するが、中期頃を境に激減し、これと同時に卵巣内の含量が急増する(第7図)。雄にはこのような蛹初期の急増も中期以後の激減もみられず、雌においても、卵巣を摘出しておくと中期以後に体液中のビタミンB2の減少は起らず、対照蚕の4倍以上もの量が蛾の体液中に含まれていた。また片側の卵巣を除去すると残った卵巣における卵数、卵重が増加し、卵内のビタミンB2含量も増加した。

第7図 蛹の体液と卵巣とにおけるビタミンB2の消長(江口(89))
◎:体液中のB2(γ%)、○:卵巣中のB2(蛹1頭当りのγ)
●:1卵巣当りのB2(mg)、横軸は化蛹後の日数
これは、一見、ビタミンB2が亜鉛その他と共に、卵の発育に必要な栄養物質の一つではないかと思わせる結果であるが、od油蚕(第Ⅰ連関群の遺伝子)における結果と考え合わせると、問題がなお残っている。od油蚕の蛹期におけるビタミンB2の消長は、その曲線の形に関する限りは、第7図に示した正常蚕の場合と違わないが、最大値を示す蛹中期においても、体液中の含量は正常蚕の中期以後における激減した量と殆ど同じレベルであり(90)、卵における含量も正常の約半分である(66)。この場合、od蚕の造卵数、孵化歩合などについての記載はないが、普通、od蚕の造卵数および孵化歩合に特に顕著な欠陥のないことからみると、卵巣の発育および卵の造成に関するビタミンB2の役割を第7図の消長のままに受取ることはできない。
3・ヒドロキシキヌレニンも、後に述べるように、蛹期における卵内への移行が顕著であるが、卵の造成には関係がない。中曾根(665)は卵巣のグリコーゲンが蛹の中期から激減することをみたが、山下・長谷川(1171,1172)はこのような減少を記載していない。土井良(87)は、体液蛋白の電気泳動像を調べ、雌に特異な泳動帯FLおよびFPの存在を示した。FL(幼虫型雌蛋白)は5令雌に限って存在し、FP(蛹型雌蛋白)は化蛹後に現われ、雌蛾においても痕跡的に認められると云うが、蛹の発育に伴なうその消長は明かにされていない。
ii 卵黄および卵膜の形成 蚕卵の卵黄は脂肪性の顆粒と蛋白性の顆粒とに大別されるが(290,496,501)、町田(496,501)は脂肪性卵黄にa、bの2種を区別し、卵黄形成を次ぎのように考えた。
脂肪性卵黄aは、生で生理食塩水中に取出すとブラウン運動をする微小顆粒で、卵母細胞、栄養細胞および包卵細胞中のゴルヂ体から生じ、卵母細胞が分化すると直ちに現われ初める。脂肪性卵黄bはaよりやや大きく、それ自身はブラウン運動をしない。栄養細胞核の仁が崩壊し、これが核膜を通り、細胞質中に出て卵母細胞に流入しb顆粒を作るもので、卵母細胞と栄養細胞との位置的関係がきまってから(Ⅰ2Aa)現われる。蚕自性卵黄は、化蛹後、包卵細胞が卵母細胞を囲む1層の上皮組織を作ってから現われ、包卵細胞のミトコンドリアに由来するか、あるいはこれと何らか密接な関係を有するものであろう。
これは従来最も信頼されている研究結果ではあるが,。細胞の微細構造およびその機能についての知見の乏しかった当時の研究であるから、新しい方法と考え方によって再検討し、補足すべき問題を含んでいる。
宮およびその協力者は電子顕微鏡および組織化学的方法によって卵黄形成の研究を進めている。未だ詳細な結果は報告されていないが、栄養細胞から卵母細胞に供給される脂質は初期だけに限られ、大部分は包卵細胞から来るらしく、蚕自性卵黄も包卵細胞から流入する物質によって作られるらしい(558,559,563)。栄養細胞において、核膜を通しての核と細胞質との連絡は認められるが、核からの逸出物が脂肪性卵黄に転化する像は観察されなかった(557)。
大槻(774)は卵黄形成の種々な時期に蚕卵を遠心して卵の内容を分画し、その各層の量および組織化学的性質を調べる方法によって、正常卵と包卵被膜細胞の遺伝的に退化する系統の卵とにおける卵黄形成を比較した結果、栄養細胞から卵母細胞に流入する物質は、脂肪性のものも若干含んではいるが大部分は好塩基性蛋白質の反応を示す物質で、このground
substance(基礎物質)に包卵細胞から来る物質が働いて蚕自性卵黄を形成するものと考えている。岩下(294)も、蛋白性卵黄は栄養細胞と包卵細胞とから流入する好塩基性物質によって形成されるらしいと云う。
江口ら(91)は蛹卵巣の組織蛋白を蒸溜水で抽出し、その電気泳動像において、中期には殆ど観察されない1本のバンドが後期には明瞭に認められるようになることを報告した。上田(1090)によれば、卵巣のチトクローム・オキシダーゼは、ナジ反応によっては栄養細胞にだけ認められ、その卵巣生体重当りの活性度は蛹令と共に低下し、栄養細胞の機能と密接な関係かあるらしい。
卵黄形成が終ると卵の外側に卵膜が形成される。蚕の成熟卵の卵膜は卵母細胞を直接に包む卵黄膜とその外側を覆っている卵殻とであるが(第23図)、町田(496,501)によれば、卵黄膜は卵母細胞の周辺細胞質から形成され、卵殻は包卵被膜細胞から分泌される。町田は卵殻を、3層からなる内卵殻と1層の外卵殻との4層に区分しているが、層の区分は研究者によってかなり相違している(27,28,29,30,524a,568,783,916)。
化蛾の3日ぐらい前になると、卵管下部の卵は包卵被膜から離脱して下降し初める(第4表)。
c 蛾
化蛾の前後には、卵の母体内での成熟最終段階である包卵被膜離脱が急速に進む(662,952)。清水・堀内(852)によると、その進み具合は蚕の品種によって特徴があり(第7表)、大造および輪月では化蛾前夜の離脱が非常に多いが、他の品種においてはそれ以前から徐々に離脱が起こっていて、化蛾前夜には少なかった。交尾蛾と末交尾蛾とを化蛾当日の午後4時に比べると、交尾蛾においても予期したほどには離脱が促進されていなかった。しかし、化蛾当日午後4時の末離脱卵歩合は品種間で大差がないことをみると(第7表)、このときの末離脱卵は卵形成に関する何らか共通的な原因によって特に成熟のおくれた卵であり、そのために交尾の影響が現われにくいのかも知れない。また、化蛾当日午後4時および化蛾翌日午前9時の調査において産卵の多かった蛾は、産卵の少なかったものに比べて離脱率が幾分高かったと云うが、これは卵の成熟の進んだ蛾が早く産卵すると云うことに外ならないと思われる。
清水らによれば(852)、温度と包卵被膜離脱との関係は品種によって相違する。大造においては15℃での離脱が緩慢で、20℃、25℃、30℃と温度の上昇するに連れて直線的に離脱が進むのに対し、日115号においては、15℃でもかなり離脱は進むが、25℃で増加が止まり、30℃では25℃よりも却って離脱が少なくなって、これらの品種の消化率と温度との関係に似ていると云う。沓掛(471)は産卵歩合と包卵被膜離脱率との関係を調べている(Ⅳ3Ca)。
卵は、普通、卵母細胞第一成熟分裂の後期初期の状態で産み出される(814)。
| 調査年次 | 蚕品種 | 化蛾前日 午後4時 |
化蛾当日 | 化蛾翌日 午前9時 |
|
| 午前6時 | 午後4時 | ||||
| 1937 | 欧18号 欧16号 小石丸 輪 月 |
45% 38 29 43 |
41% 28 23 21 |
26% 22 16 14 |
14% 11 7 8 |
| 1949 | 欧18号 日115号 支105号 支110号 大 造 輪 月 |
43 30 35 31 61 53 |
25 21 32 30 37 29 |
27 16 20 16 19 17 |
18 8 13 8 6 5 |
B その他の器官
a 内部生殖器官
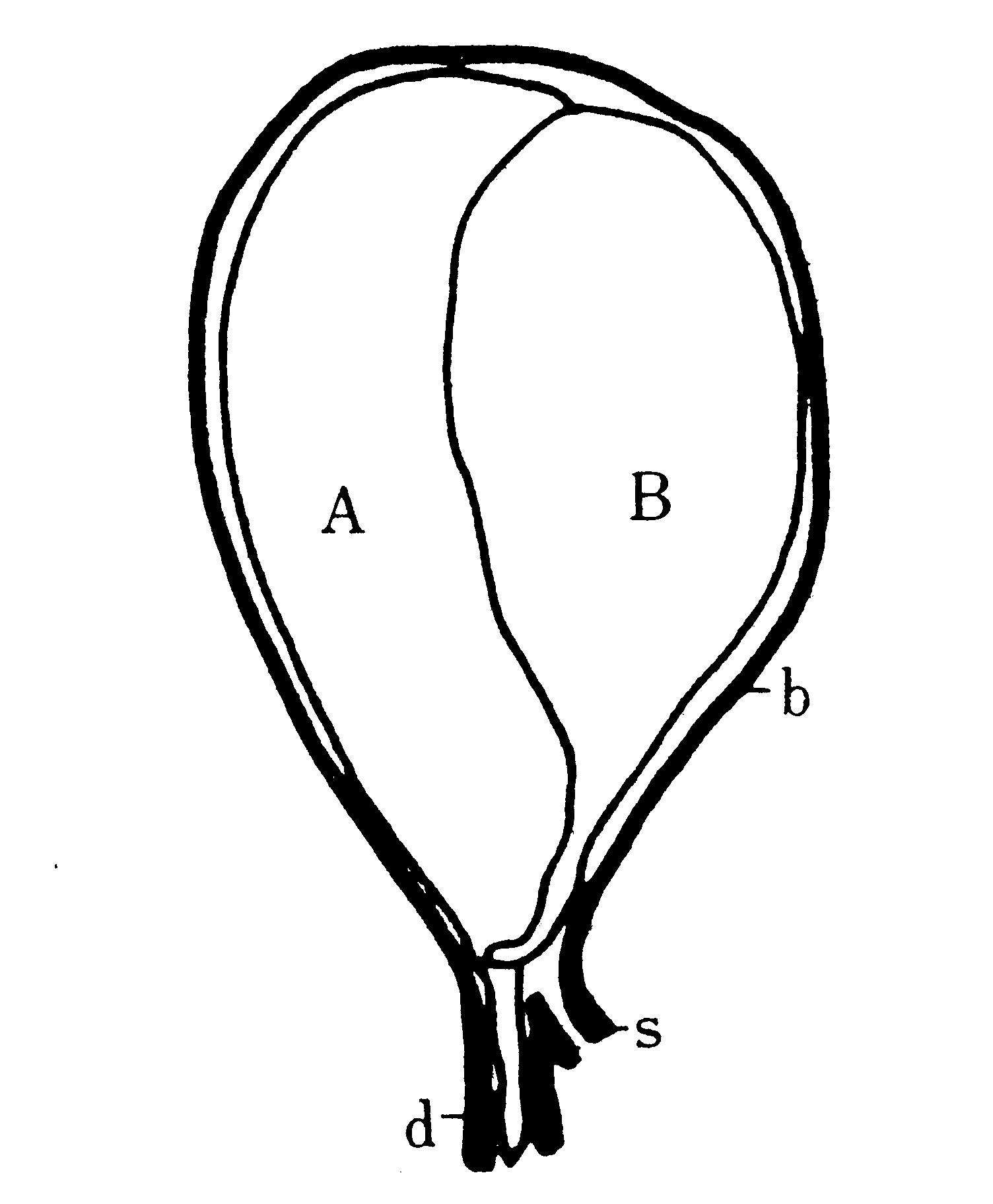
幼虫の生殖器官は第8節背面の皮膚下にある卵巣と、それから出て後方にのび、第10節後端の皮膚に付着する紐体(第2図)、第11節の石渡前腺および第12節の石渡後腺(第19図)とである。紐体は導管とも呼ばれるが、幼虫期には未だ管状構造ではないので、導管とは云い得ない。蛹になってから初めて管状となり、石渡前腺に由来する輸卵管と接合するが、紐体に由来する輸卵管枝と輸卵管との境が貫通するのは化蛾の前後である。石渡腺は、腺と云う名前はついているが分泌腺ではなく、前腺からは輸卵管、前庭、交尾嚢、受精嚢、交尾嚢導管、精子管および螺旋管(第8図)が形成せられ、後腺からは膣および粘液腺ができる(792,1096)。
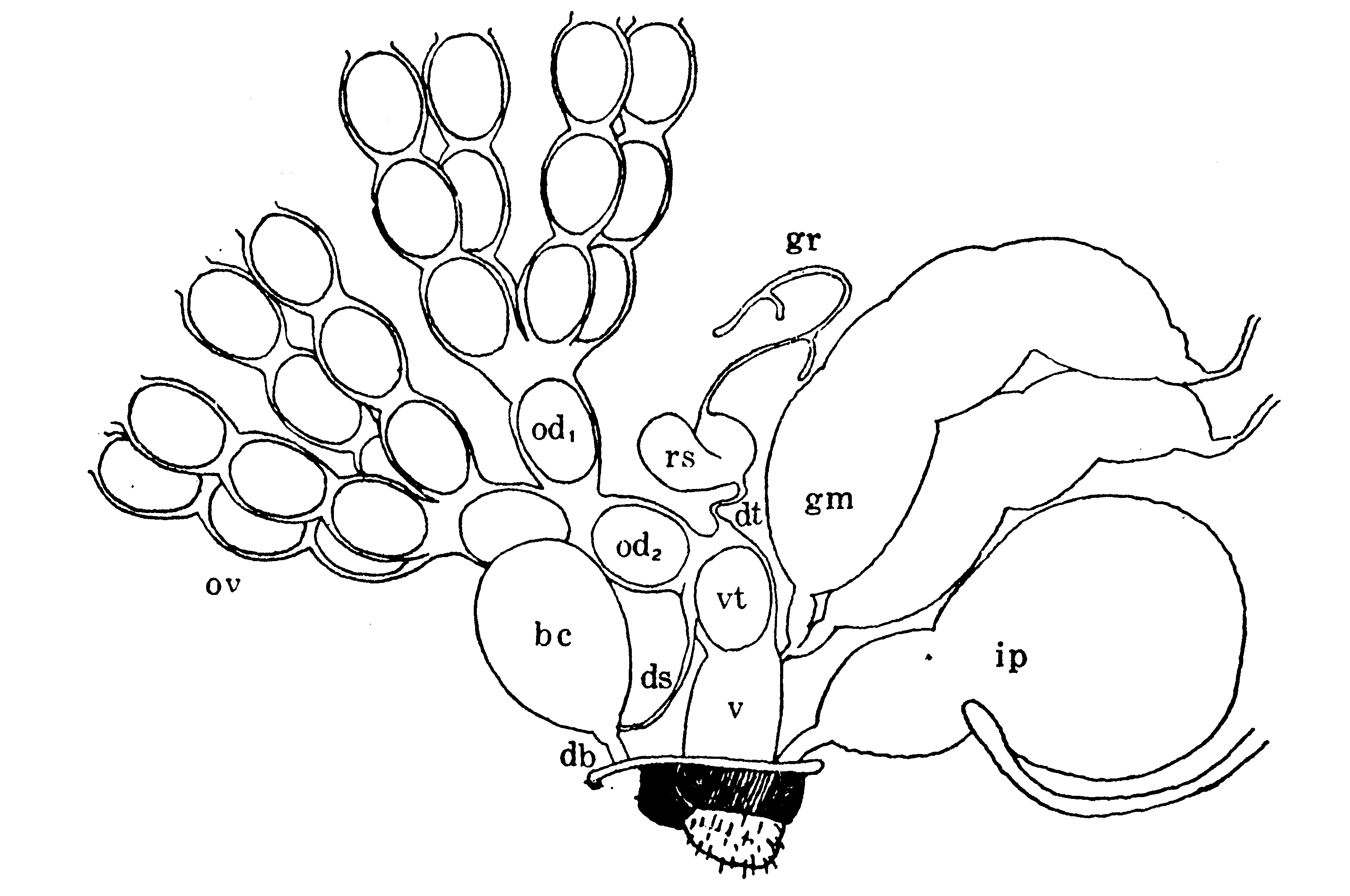
第8図 雌蛾の内部生殖器官(模式)(Omura(759))
bc:交尾嚢、db:交尾嚢導管、ds:精子管、dt:螺旋管、gr:受精嚢付属腺、gm:粘液腺
ip:後腸、od1:輸卵管支、od2:輸卵管、ov:卵管、rs:受精嚢、v:膣、vt:前庭
交尾嚢導管(大村は初め(746)交尾嚢管と呼んだが、後(1117)、交尾嚢導管に変更した)は交尾の際に雄の陰茎の挿入される部分である。精莢(Ⅰ3C)に包まれて交尾嚢内に射出された精子は、白身の運動と管壁の蠕動とによって精子管を通り、一旦膣に出た後、螺旋管を上って受精嚢に入る。螺旋管の内部には更に小さなキチン質の受精小管がある。精子は、受精の際には、この小管を通って自力で下降し、丁度下降して来る卵が前庭隆起に支えられて卵門を受精小管の開ロ部に向けたときに(第8図)、卵門を通って卵内に侵入する(746,747,759)。
卵管内の卵は卵門が上部に向くように形成されており、そのままの姿勢で輸卵管を下ってくるので、精子の侵入に都合がよい。1蛾採りの卵を調べると、卵が総べて蛾のいた中心部へ卵門を向けて産み着けられているるのはこのためであるが、稀に逆位のものがあり、逆位のものには不受精卵が多い(569,953)。このような不受精卵は、卵管を下り輸卵管を通る間に何らかの原因によって逆位になり、精子の侵入に不都合な姿勢で前庭を通過したものと考えられるが、実際に、藤本(97)は、遺伝的に卵管内での卵の逆転が起こり易く、そのために不受精卵の多発する系統のあることを報告している(Ⅰ2C)。逆位卵は蛹(953)および蛾(99)の冷蔵によっても増加する。逆位の起こるのは卵が完成し、包卵被膜を脱いでから後が普通であるが、稀には包卵被膜から離脱する前に逆転するものがあると云う(997)。
逆位卵が不受精卵になり易いことからみて、精子が自身で運動し得る範囲は広くないように考えられるが、松永(522)は、産卵後自然死した雌蛾の体内から着色した卵を取出し、翌春まで保護した後孵化させ、飼育して、その幼虫およびそれの作る繭が、交配に用いた雄蛾からの性質を表わしていることを認め、また、交尾後に産卵孔を焼灼して産卵不能にした雌蛾を、自然死した後に解剖し、その卵管、輸卵管および膣内に受精卵のあることを確かめ、精子は卵管内に上って卵を受精させ得ることを明かにした。この場合、受精卵か否かは、飼育して遺伝形質の発現を調べて判定した。
滝沢(998)も産卵孔焼灼実験を行なってこれを追認した。産卵孔を焼灼した蛾においては、膣が破裂して腹腔内に卵の出ていることが多いから、焼灼の衝撃または卵の下降する圧力によって精子が機械的に卵管内に押上げられたり、吸上げられる可能性も考えられないことはない。
正常の産卵では5.3~10.0秒に1個と云う速さで卵が産み出されるので(759)、逆位で前庭を通過する卵には精子の侵入する余裕がないが、時間をかければ精子は案外遠くまで運動し得るのかも知れない。長谷川(152)は、十分な時間交尾させた雌においては、最初に産み出される卵が未だ前庭まで下っていないときに既にその下部に精子が付着していること、およびこのような雌を産卵させた後に解剖して調べると、輸卵管まで上っている精子が産卵前のものにおけるよりも多いことなどを報告している。
前庭を通過した卵は、膣に開口している粘液腺からの膠着物質を、主に卵の裏側(台紙に付着している側)にだけ付与されて産み出される(842,843,962)。粘液腺の細胞組織学的研究には鈴木、辻田ら(891,1081)、膠着性の遺伝については鈴木・平田(888)の報告がある。
交尾した雌蛾が未交尾蛾に比べて早く産卵することからみて、交尾は産卵を誘発する刺戟になるものと考えられるが、大村(748)は、人工助精、去勢雄(精莢を射出するが、その中に精子を含んでいない)との交配、および三倍性雄(精子は受精嚢にまで達して盛んに運動しているが受精小管に下ることがないと云う)との交配など種々な場合の産卵状態を調べた結果から、精子のない場合の交配は勿論、精子があっても、それが受精嚢に入るまでの運動は産卵誘発刺戟として無効かあるいは不十分で、刺戟として有効なのは、受精嚢から受精小管を下って前庭に達する間の精子の行動であると結論した。しかし、長時間に亘って交尾している雌蛾を解剖して、前庭およびそれより下方にある卵を調べると、既に精子の侵入している卵があることからみて、精子の行動が直ちに産卵を誘発する訳ではなく、従って産卵を単なる反射運動と考えることはできないと云う。
福田(107)は、生理食塩水中での卵管および輸卵管の蠕動の実験によって、産卵を起こさせる直接の要因は蠕動であるとし、精子が雌の生殖系統内のある部分に侵入すると、蠕動誘発物質(末交尾雌および雄蛾の体内にも若干ある)を増産するか、既存物質を賦活するか、あるいは精子の侵入によって起こるある種の体液変化が同様の働きをするか、または卵管の感受性を高めるなどのことが考えられるとしたが、川瀬(365,366)は、生理食塩水中に取出した卵管の蠕動を誘発する物質は特別なものではなく、Ca++イオンに外ならないと報告している。
藤本(99)によれば、種々な品種の雌蛾を生理食塩水中で解剖し、液温5℃、7.5℃および10℃において観察すると、卵管の蠕動はこれらの品種の蛾の冷蔵中における産卵(Ⅳ4A)と符合する動き方を示し、産卵の多い品種の卵管はよく動くと云う。
室賀(612)は、腹部第4神径節を摘出すると、交尾しても産卵せず、脳とこの神経節との連絡を断つと受精しても産卵数の少ないことから、腹部第4神経節は産卵機能に直接関係し、脳は産卵に関する何らかの指令をこの神経節に与えているのであろうと考えた。山崎ら(676,677,678,1176,1177)も同様な実験を行ない、また明暗が産卵に影響する(Ⅳ5A b)のは光が複眼を通して作用するのであるとし、雌蛾に産卵を起こさせる直接の原動力は頭部内神経節で、これが第4複節神経節と協同して働くのであると結論した。福田(113)も最終神経節(腹部第4神経節)が産卵に関係することを認め、上に述べた体液的要因と神経系の活動とが相まって盛んな産卵を誘発するものと考えている。蚕の産卵は暗いと促進されるが、脳または食道下神経節を摘出するとこの反応が正常でなくなると云う(428)。
b 外部生殖器官
雌蛾の外部生殖器官を第9図によって説明すると(492,1036)次ぎの通りである。側唇は他の昆虫の産卵管に相当するもので、その腹端に産卵孔が開口する。交尾孔は交尾嚢導管(第8図)の開口部である。側唇の左右の鋸歯板との間の部分は嚢状に膨出して側胞となる部分で、雄を誘引する物資を出す。性誘引物質については竹田(971,972,974,976)の研究があるが、Butenandtらによって化学構造が決定された。家蚕の性誘引物質には桑蚕の雄蛾が反応し、桑蚕の性誘引物質には家蚕の雄も反応する(755,974)。その分泌は明るいと幾分抑止されるが、完全には停止しない(979)。
側唇は幼虫の尾脚、誘引腺は同第8腹節の後方約3/4と第9腹節の1部に亘る腹面(左右は気門下線まで)の範囲、産卵孔は同じく左右の尾脚間の皮膚からそれぞれ作られる(973,975,980,981)。
第8腹節腹板の後縁中央に彎入部があり、その左右には鋸歯状の切れ込みがある。この彎入部は、交尾の際に雄蛾の鈎器および竜骨(第13図)に挟まれて連結器の役目をする。
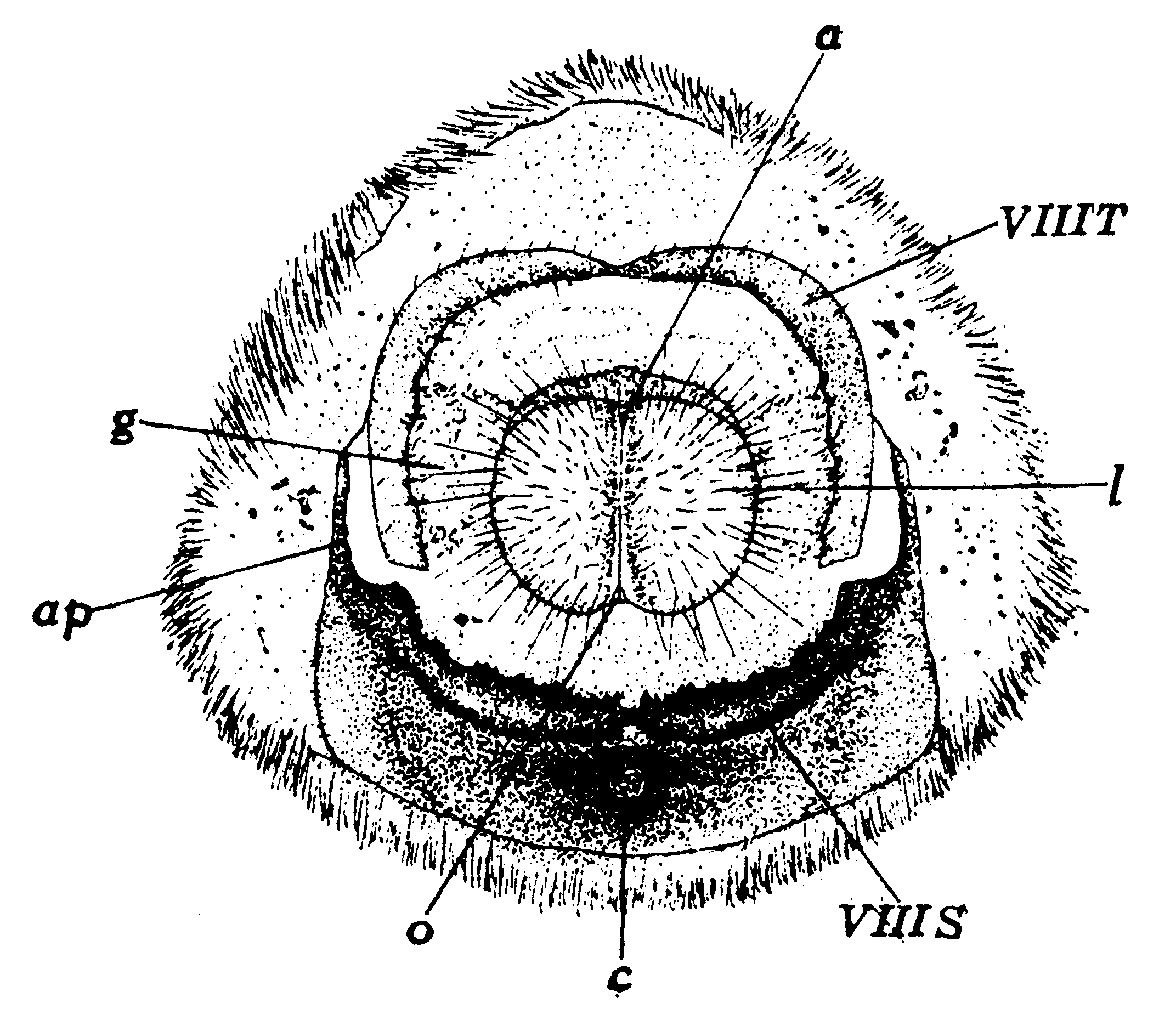
第9図 雌蛾の外部生殖器官(田中(1036))
a:肛門、ap:鋸歯板棒状突起、c:交尾孔、g:側胞として膨大突出すべき部分
l:側唇、o:産卵孔、ⅧT:第8腹節背板、ⅧS:同腹板
C 雌生殖器官の異常
畸形あるいは異常として報告されているものの形態は極めて雑多であり、また遠心力とか放射線とかを作用させて起こした異常を数えあげれば限りがない。ここでは蚕種製造に関連の深い畸形を3群に大別して、数例を述べる。
a 正常型の個体変異
解剖してみると、左右の卵巣の大きさや造卵数が相違している(219,840,841)とか、同じ第8節内ではあるが、左右の卵巣が正しく向き合わないで、位置がずれている(1004)と云うような変異はむしろ正常な姿である。
b 遺伝的な異常
遺伝学や発生学の研究材料に用いられている遺伝的畸形には種々なものがあるが、実用品種にはこのようなものの入らないように注意が払われている。しかし、時々問題になる場合があり、最近の例としては支115号の雌雄モザイクがこれである(184,185)。雌雄モザイクと云うのは、1頭の蚕の体に雌性の部分と雄性の部分とが混り合っているもので、その程度や発現部位によっても異なるが、交尾、産卵の正常に行なわれない場合が多い。遺伝的なものであるから、支115号の系統分離によって育成された晴と云う品種にもこれが発現した。但し、これらの系統の総べての個体がこの素質を持っていると云う訳ではなく、劣性の遺伝子を持つ個体が僅に含まれているために時々発現するのであるから、系統内に広く分布してはいるらしいが、実害は殆どないと云われている。これらの系統から無選択に採った雌にod油の雄を交配してF1を調査した結果によれば、支115号2,707頭中に25頭、晴441頭中に6頭の雌雄モザイク蚕が発見された。od油は劣性の伴性遺伝子で、上記のように交配すると、異常がなければ、F1雄は全部正常皮膚、F1雌は全部油性を現わす筈であり、雌雄モザイク蚕は正常皮膚と油とのモザイクとして現われるから発見が容易である。遺伝学の材料として有名なモザイク遺伝子は勝木モザイクと呼ばれているものであるが、支115号の雌雄モザイクはこれとは異なる遺伝子によると云う。支115号からは三倍体個体の出ることも認められたが、その成因は根本的には雌雄モザイクの場合と異ならない。即ち、雌雄モザイクは、成熟分裂の異常によって卵内にZ染色体を持った核とW染色体を持った核との2個の卵核ができ、これが別々に受精してそれぞれに発育する結果、雌雄の性質の混り合った蚕ができるのであるが、三倍体は、上記の2卵核が別々でなく、一緒になって1個の精子により受精してできるものと考えられる。
古い品種であるが、日110号に不産卵蛾が出て、解剖してみると、膣と輸卵管との連絡不完全、膣の縊れなどの欠陥があるため、卵巣には卵が形成されており、交尾もするが、産卵し得ないことがわかった。その原因としては、飼育環境や成虫盤の異常など種々な考察が行なわれた(497,886,955)。最近、坂之下(792)は、過剰半月紋を有する遺伝的に輸卵管と輸卵管支との結合しない系統の蚕について、紐体末端分枝の伸び方の異常がその欠陥の原因であると説明した。
大宮(739)によれば、日122号、日124号、支122号(大)、支124号などの正常産卵蛾を、産卵後4-5日たってから解剖してみると、卵管内に配列の異常な卵や大形の未完成卵があったり、卵管と輸卵管との接合部に数個の卵が詰ったりしていて、それより上部の卵の下降を妨げているようにみえる場合や、卵管の1部が破れて卵が体腔内に出ている場合が非常に多かった(35頭中にこのような異常を示した蛾が28頭もあった例がある)。産卵後、日のたった蛾であるから、卵管が崩壊し初めたための異常を含んでいるとは考えられるが、このような異常個体においては、正常産卵蛾ではあっても、異常のないものに比べて体内残留卵歩合が高いと云うことは注目される。
既に述べた藤本の不受精卵の多い系統(Ⅰ2Ba)は支105号に由来するもので、逆位卵が生ずるのは卵管の蠕動が異常に激しいためと考えられている。
遺伝学の材料になっている各種の畸形は実用に縁遠いもののように考えられ勝ちであるが、例えば、異常生殖腺蚕(Ⅰ2Abi)の研究は造卵数の問題についての考察に重要な意味を持っている。直接の実用問題ではないが、市川(268)の発見したENと云う突然変異は卵巣の発生についての貴重な研究材料になった。ENホモの蚕は催青死卵になるが、その胚は第1~10節の全部に胸肢様の付属肢を有し、正常の腹肢を欠いている。宮(562)の研究によれば、この胚の生殖細胞は正常に分化し、増殖するが、腹部に生殖隆起が形成されないため、生殖細胞かそれに包含されることもなく、体腔内に取残されて生殖腺を形成しない。即ち、生殖腺の形成には生殖細胞と生殖隆起との二つの要素が必要で、この何れが欠けても完全な卵巣あるいは睾丸はできないのである(Ⅰ1)。
包卵被膜異常の系統が卵黄形成の研究に役立っていることも頂に述べた(Ⅰ2Abii)。
c 偶発的な異常
卵管数が正常の8本より少ない蚕、あるいは多い蚕が縷々発見される。卵管数の少ない個体は造卵数も少ないが、実害のあるほどの頻度ではなく、少数蚕卵蛾の発現とも関係はないと云う(745)。清水・堀内(853)は、このような蚕の出現は春蚕期よりも夏蚕期に多いから、人工孵化の塩酸刺戟の影響ではないかと考えている。ただ、遺伝的な素質のあるものは、種々な外因が作用すると異常を発現し易いから、ここで云う遺伝的か偶発的かの区別は便宜上のものに過ぎない。卵管の数が1卵巣に普通4本であると云うことは、発生の過程において、第6-9節に生じた生殖隆起が連絡、収縮して生殖腺を作ること(Ⅰ1)と関係がありそうに思われるが、宮(562)はこの関係はないものと考えている。
市川(269)は、第8節腹側に突起のある蚕を発見したが、この突起には腹肢と気門とが1対ずつついており、解剖してみると、その中に余分の遊離した小卵巣が含まれていた。これは生殖腺の異常と云うよりは重複胚の形成で、発生学的に興味がある。これに類似した異常の出易い系統も知られている。佐藤・田中(820)は人為単為発生によって過剰卵巣蚕の出現したことを報告した。三谷・渡会
は不産卵蛾を解剖して、内部生殖器官の種々な異常を記載している。
3 雄の生殖器官
A 睾丸
a 睾丸の発育および精子の形成
幼虫の睾丸は、卵巣と同じく第8節背面、星状紋のある部分の皮膚下に、背脈管を挾んで左右に1対あるが、内令の卵巣に比べて大きく、丸みがあり、腎臓形を呈し、その凹面で相対している(第10図)。
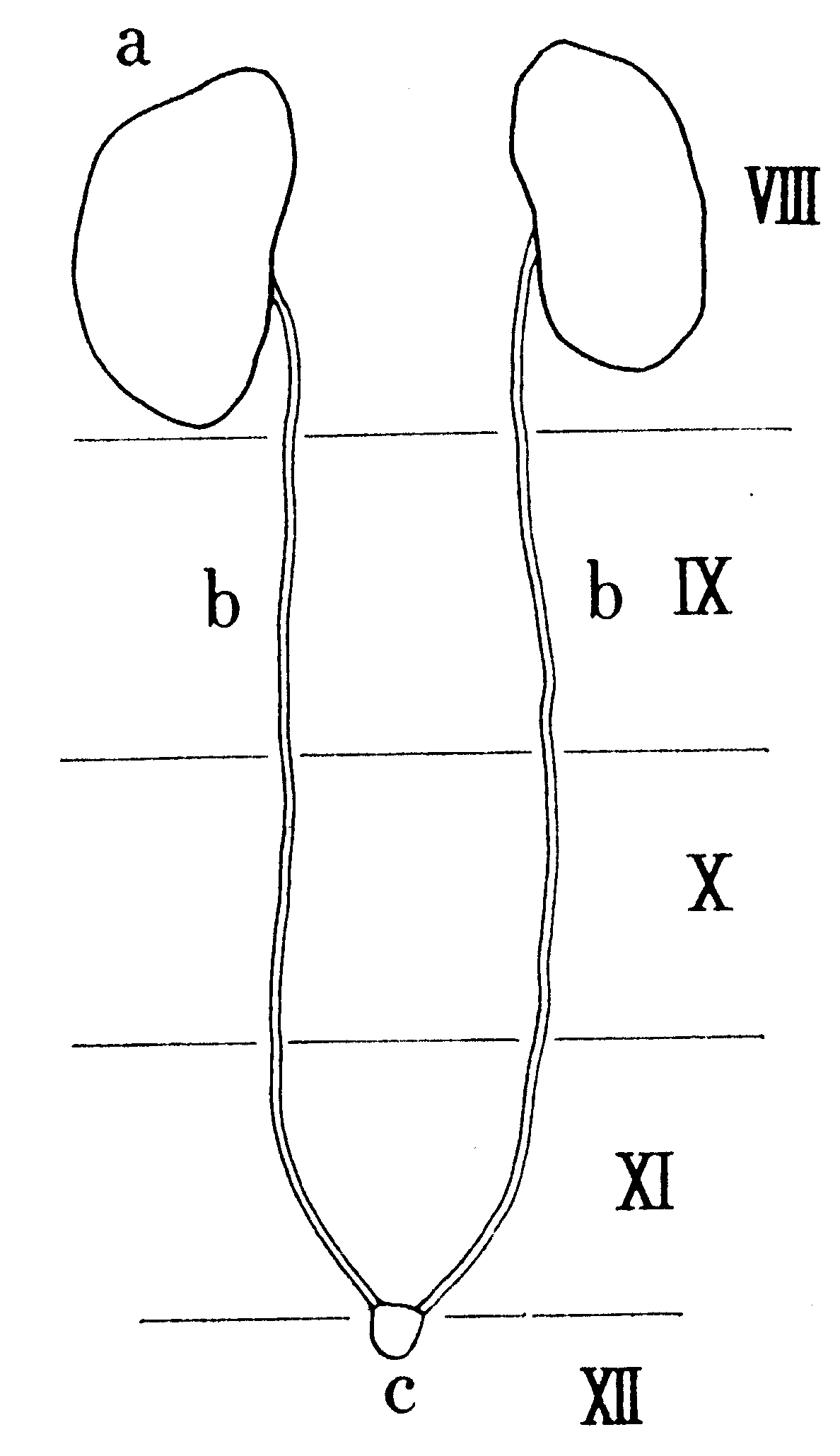
第10図 幼虫睾丸の位置(模式)
a:睾丸、b:紐体、c:ヘロルド腺、Ⅷ-XII環節
(背面からみた位置。田中(1035)によって描く)
孵化当初には内部の区画が明瞭でないが、間もなく隔壁が発達して、普通、4個の精室に分かれる(第11図)。
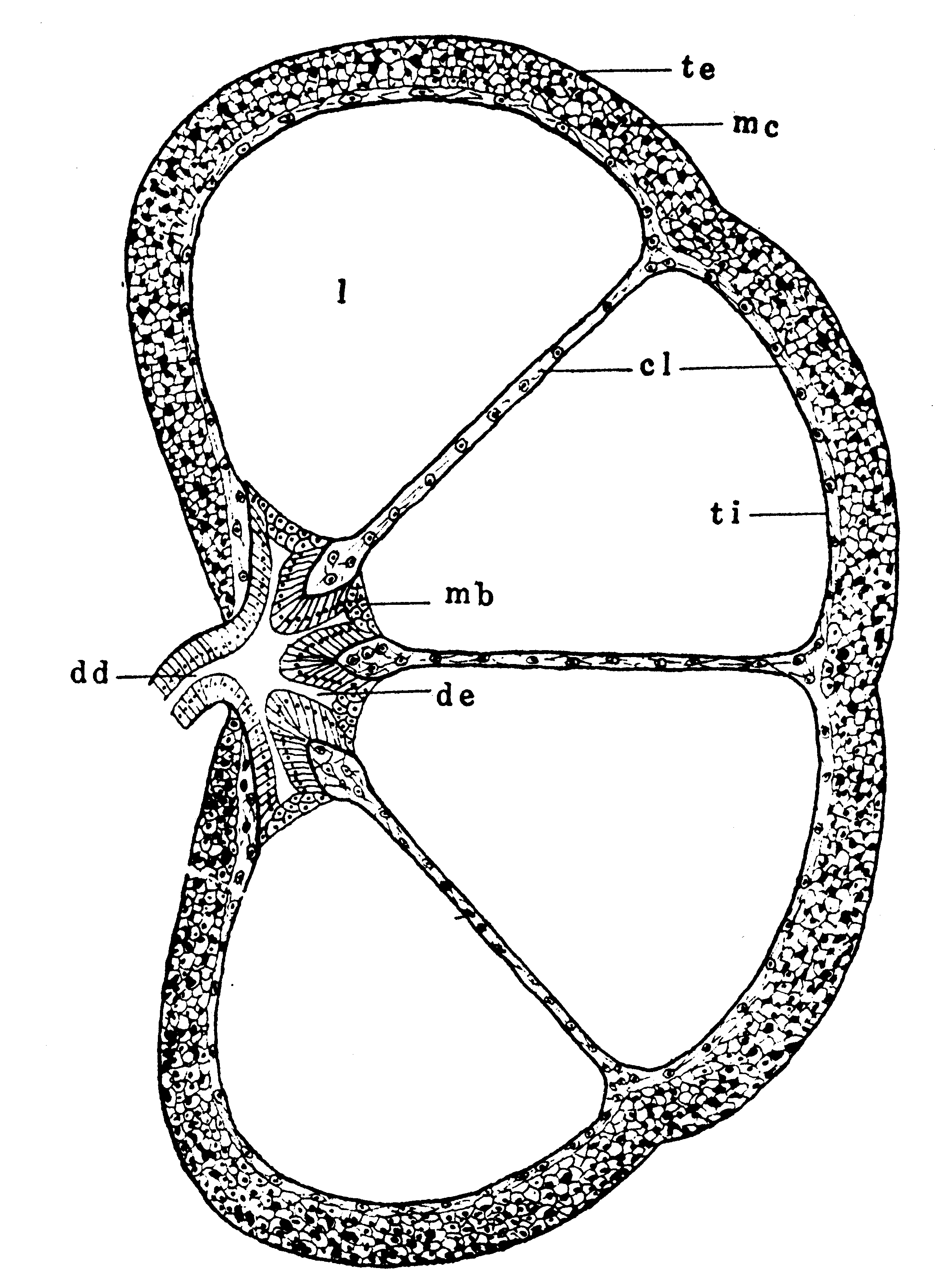
第11図 幼虫睾丸の縦断(模式)(Omura(758))
cl:精巣被膜、dd:輸精管、de:小輸精管、l:精室
mc:共通膜、mb:基底膜、te:外膜、ti:内膜
精室の頂端部には尖端細胞と呼ばれる大形の細胞がある。その起原や機能については古くから種々な見解があるが、最近では、宮(554)が、孵化直前の胚の生殖腺において尖端細胞と考えられる細胞を観察し、生殖細胞の変化したものであろうと考え、須貝(867)は、精巣被膜組織から分化するものとして、町田(490)の考えを支持している。尖端細胞は卵巣においては特別の機能を持たないものと考えられているが、睾丸では活溌な活動を示し、第1眠期頃になると、この細胞に回って多数の精原細胞が尾状の突起を出し、同心円状に配列するのがみられる(490,494,500)。須貝は、このときの尖端細胞に多糖類(グリコーゲンと考えられる)の反応を示す顆粒の蓄積が始まり、尖端細胞およびこれを取巻く精原細胞にアルカリ性および酸性フオスファターゼの活性が強いことを観察し、尖端細胞が精原細胞に対する栄養供給細胞であることを実証するものと考えている。
須貝によると、尖端細胞を取囲むそれぞれの精原細胞が分裂して2個になると、一つは将来精母細胞として発達するもの、他はこれを取囲む皮嚢細胞に分化する。
精母細胞に分化するものは、その後6回の分裂を行ない、26=64個になったときに分裂を止め、大きさを増し、第一次精母細胞に分化する。これが更に2回の成熟分裂を行なうので、1個の皮嚢内には256個の精子細胞ができ、これが1個の精子束になるものと計算される(334)。この間の精子形成の経過と蚕令との関係は、佐渡(776,779)によれば第8表の通りである。種々な時期の幼虫にH3-チミジンを注射し、標識された精子の形成を調べた結果から考えても、第一次精母細胞におけるDNA合成期の最後から精子完成までが18日、精原細胞期は約7日で、第8表とよく一致している(778)。
| 孵化後の日数 | 蚕 令 | 最も発育の進んだ性細胞 |
| 6日 10-11 16 17-18 18-19 24 |
Ⅱ令 2日目 Ⅲ令 2日目 Ⅳ令 3日目 4眠-Ⅴ令1日目 Ⅴ令 2日目 Ⅴ令 6日目 |
第一次精母細胞 合糸期 〃 太糸期 〃 収縮期および分散期 〃 移動期および第一成熟分裂中期 第二次精母細胞および若い精子細胞 完成精子 |
滝沢・玉沢(1010)によれば、成熟分裂は5令餉食後15時間目頃から始まり、精子束の現われ初めるのは餉食後21時間目頃であるが、1日のうちでO-3時と12-15時との2回、特に分裂の盛んな時刻がある。
1睾丸内で作られる精子の数については町田(493)の調査がある。日107号につき、1睾丸内の完成精子の数をかぞえると、化蛹14日目に化蛾したものについて云えば、完成精子は熟蚕前日から一部の睾丸に現われ、熟蚕では総ての睾丸において認められ、以後増加を続け、化蛹9日目に最大数に達し、その後11日目までは殆ど増減がなかったが、12日目には減少を示し、以後は急激に減少し
た。化蛹9日目には、無核精子になるべきもの以外には未完成精子はなかった。
12日目には完成精子が睾丸から出初め、導管、貯精嚢などの中に精子が認められた。増減のなかった第9-11日目に観察された有核精子束数の平均を数学的に補正すると、補正に用いた定数Kの値のとり方によって、1睾丸内の精子束数は2,936.5-3,915.3個と計算される。
30個の精子束につき、その中に含まれる精子数をかぞえると、川口の計算したような256個のものは唯1例だけで、他は211-251個の範囲の数を示し、平均は234.0個であったから、形成の途中で退化するものがあると考えられる。町田は、これらの精子束数平均および精子束内精子数平均から、1頭の雄の生産する有核精子数を1,400,000-2,000,000個と計算した。佐渡(777)も、1睾丸当りの精子束数を、上記に近い2,500個と計算している。
卵巣の重量が蚕の発育に連れて増加するのとは異なり、睾丸の重量には発育時期によって増減がある。江竜(95)によれば、化蛹直後を100とする睾丸重量の指数は、熟蚕期51-50、化蛹前日91-97、蛹中期61-70、化蛾前日50-55、化蛾直後44-38で、化蛹直後が最も重い。この増減の原因は共通被膜内の脂肪の蓄積および消費、睾丸内からの分泌物の流出、精子の下降などによって説明されている。
b 無核精子および精子の長さ
無核精子と云うのは、他の鱗題目昆虫においても知られている受精機能を欠いた異常精子であるが、正常の有核精子に混って普通に形成される。町田(499)によれば、幼虫期に成熟分裂を完了する精母細胞は全部有核精子となり、蛹期の後半になってからの成熟分裂は全部無核精子を形成するもので、無核精子は精室の基部までは下降するが、そこで退化、消失して睾丸の外に出ることはない。大村も初めは(746)、無核精子は運動力がなく、睾丸から脱出し得ないと記載したが、その後、稀に輸精管膨大部内にも含まれることを観察し(758)、更に遺伝的精子欠乏症の蚕においては、交尾によって雌の受精嚢内に移行する無核精子のあることを認めた(890)。入来(247)は、無核精子は睾丸外に出るばかりでなく、射精によって交尾嚢に入り,、更に自力で受精嚢へ移行すると述べている。勝野(322)は、精爽腺の分泌物に原因があると考えられる“ちぢれ精子”が雌の交尾嚢内に移行していることを報告したが、その後、これは無核精子が精莢腺の分泌物によって縮れることがわかったと云う。しかし、精莢腺以外にも原因があるらしい(私信による)。
無核精子の行動についての観察結果にこのような相違のあるのは、形態だけからみて無核精子と呼んでいるものに、性質の異なった異常精子が含まれているためかも知れない。
無核粒子は、精子形成に必要な栄養に関して何らかの欠陥があるために生ずるものと考えられているが、睾丸移植実験の結果からは明確な結論を下すことができなかった(499)。
須貝(864,865,866,868,869)は、精子の形成過程を組織化学的に研究した結果から、精母細胞に対する栄養供給が不良になると、その程度に応じて、無核精子の形成、精母細胞あるいは精原細胞の退化がおこるのであろうと推論しているが、実際に絶食試験を行なってみると、5令起蚕に3回給桑した後に絶食させ、絶食時間が48時間以上になると精巣被膜組織中のグリコーゲンが著しく減少した。この減少は絶食を止めると回復するが、熟蚕期になって睾丸を調べてみると、異常形態を示す精子束(退化型精子束)が形成されており、正常の場合には幼虫期の睾丸には有核精子束のみが認められるのに対し、48時間絶食の後、更に極度の給桑制限を続けたものや絶食72時間(以後は正常に給桑)のものにおいては、僅かながら無核精子の形成も認められた。退化型精子束はRNAの染色性が無核精子に似ていると云う。
無核精子については、この外に、有核精子束と無核精子束との数の比(有/無)が季節によって変化し、その値の大きい品種は有核精子束が長く、原種に比べて交雑種の値が大きく、暗所飼育よりも明所(普通)飼育において大きいが(Ⅱ2Bb)、この場合の光の影響は単眼を通して作用するらしいこと、などの一巡の報告がある(195,197,198,248)。但し、これは化蛾直後のものの睾丸についての調査であって、精子は大半睾丸から出ている筈であるから、残ったものだけをみている訳で(Ⅰ3Ba)、もし有核精子と無核精子とで睾丸脱出に相違があれば、それによっても比の値が違うのではないかと思われる。
無核精子の多少が有核精子の長さに関係のあるらしいと云うことは、有核精子の長さが栄養条件によって変化することを示すように思われる。
大村(750)は、内的または外的条件によって精子形成が影響を受けた場合の異常は、精子束の長さと精子の回旋とに現われると考えている。回旋と云うのは、精子が睾丸から出ると固有の螺旋形を呈することであって、睾丸からの脱出に件なう成熟によって現われる性質と考えられている。
輸精管膨大部および貯精嚢からとった精子束の長さは蚕品種によって相違し、体の大形な品種、卵の大きな品種においては概して長いが、同一品種内においては、上記の予想に反して、栄養の良否や生殖器官の大きさなどによって影響せられず、卵の場合とは異なり、緑蚕上蔟や4令期の片側睾丸摘出などによっても長さには変化を生じなかったと云う(750)。また、支那種錫元から偶発した二眠蚕の精子束は80束平均705±2.6μ、同一蛾区の四眠蚕においては160束平均718±1.9μで、両者の間に大差がなかった。蛹を冷蔵すると精子束が短かくなるが(第9表)、マイエラ・ゼブラ×日本種のF1においては、化蛹2日、5日、10日目からそれぞれ5、10、15日間冷蔵したが、無冷蔵のものに比べて短くならなかった。但し、これが雑種強勢によるものか否かは不明である。
| 蚕品種 | 冷 蔵 (5℃) | 調査個体数 | 調査精子束数 | 精子束の長さ |
| 赤 熟 | 無冷蔵 化蛹2日目から15日間 |
10 4 |
190 71 |
687μ 630 |
| 日106号 | 無冷蔵 化蛹2日目から15日間 |
6 7 |
156 377 |
659 602 |
大村の成績には、マイエラ・ゼブラに交配した日本種の測定値がないので、原種と雑種との精子束の長さの相違が明かでないが、雑種はマイエラ・ゼブラよりは遙に長かった。この点についての平野(196)の調査によれば(第10表)、交雑種は両親品種の何れよりも長く、四元雑種は交雑原種よりは短かいが原種よりは長く、興味のある結果を示している。栄養や緑蚕上蔟などによっては影響されることのないと云う精子束の長さが、原種と交雑種との聞でこのようにはっきり相違することは、交雑種の生殖生理を考える上から注目に値する。
竹内(995)は、睾丸およびアラタ体の移植実験の結果から、前胸腺ホルモンは精子形成を促進するが、アラタ体ホルモンはこれに抑止的な作用をおよぼすものと考えている。
B その他の器官
a 内部生殖器官
幼虫の内部生殖器官は、第8節の睾丸と、これから出て後方に伸びる紐体および紐体末端の付着するヘロルド腺とからなる(第10図)。 ヘロルド腺は第12節前端腹中線に在ってその前端は少し第11節にかかっている(第19図)。紐体は幼虫期には紐状を呈し、管状になるのは蛹である。紐体からは蛾の輸精管(第12図)、ヘロルド腺からは同じく付属腺、貯精嚢、輸精管膨大部、射精管、交尾器などが形成される(237,1036,1111)。

第12図 雄蛾の内部生殖器官(模式)(Omura(760))
add:輸精管膨大部、dd:輸精管、ga:射精管中部、gl:付属腺下部、gp:射精管下部(前位腺)
gpl:付属腺上部、gs:射精管上部(精莢腺)、p:陰茎、t:睾丸、vs:貯精嚢
大村(746,758,760)によれば、睾丸内の精子が小輸精管に出るのは単なる流出によるものではなく、自力で精室の基底膜を貫き、外鞘を精室内に残し、裸になって出るのであって、束のままで残るか束が崩れるかは、基底膜を通過するときの機械的な条件によってきまる。睾丸内で活溌に運動していた精子は基底膜を通過すると運動性を失なうが、これは小輸精管および輸精管の分泌物に運動を抑止する作用があるためと考えられる。
輸精管膨大部(大村は初め貯精嚢と呼んだ)は精子を貯える部分で、その分泌液は輸精管のものに類似するが、あるいは精子に栄養を与えるものとも思われる。貯精嚢(古くから貯精嚢と呼ばれていたが、大村は初め射精嚢と名付けた)は1回分の射精量の精子を貯える部分で、その分泌液は輸精管膨大部のものに類似する。貯精嚢に連なる射精管上部(精莢腺)は精莢を作る透明、粘稠な物質を分泌する。射精管中部(白色部)の分泌物は顆粒状、白色不透明、射精管上部および下部の分泌物の接触を機械的に防止しているもののようで、射出後の精莢の先端に真珠色の小塊として付着している(第14図)。射精管下部(前位腺)の分泌液は透明、やや粘稠で、射精中の精子と混合してこれに運動性を与える。付属腺下部(乳白部)の分泌物は乳白色で、精子のあとから射出され、交尾嚢内の精子に栄養を与え、また粘稠な精液を稀釈して精子の運動を活溌にするものであろう。付属腺上部(透明部)の分泌液は透明、粘稠で、最後に射出され、交尾嚢導管内に充満凝固して交尾孔閉塞栓を形成する。
大村は、日本種一化性×支那種二化性の交雑種につき、化蛾4日前(精子は未だ睾丸から全然出ていない)、化蛾直後および化蛾4日目のものの睾丸を秤量し、その重量比が100:53:15であり、化蛾4日目のものの睾丸には正常精子は殆ど残っていなかったことから、これらの睾丸内に含まれる精子の量は、ほぼ(100-15):(53-15):(15-15)≒85:38:1で表わせるものと考えた。これによれば、化蛾の際には精子の大半が睾丸から出ていることになる。従って、睾丸からの精子の下降と交尾との間には関係がない。睾丸からの精子の下降は蛾が死ぬまで停止せずに続いている。
雄蛾は交尾しても直ちに射精するものではない。大村は、交尾すると直ぐ、相手の雌蛾を第7腹節で切断し、体内の諸組織を取除き切ロから交尾嚢を観察する方法によって、射精は交尾してから5-10分後に始まり、25-30分間継続することを確めた。
しかし、射精が始まっても直ちに精子が射出されるのではなく、最初の5-10分間は精液(精漿)だけであり、次いで5-10分間精子が射出され、そのあと再び精液だけの射出が10分間ぐらい続いて1回の射精が終る。
1回の射精が終ると貯精嚢、射精管、付属腺などは総べて内容を失い、幾分収縮するが、20分ぐらいたつと輸精管膨大部から貯精嚢へ乳白色の精液の流入するのがみられ、30-60分で殆ど充満する。射精管下部の分泌物も90分で殆ど回復するが、射精管中部は5時間、付無腺下部は10時間後になお肉眼的に認められる程度には回復せず、50時間後に第2回目の射精を行なわせたが、形成される精莢の壁は第1回のものに比べて逼かに薄かった。
勝野(320)はトーマの血球計算器を用いて精子数をかぞえ、交尾嚢および受精嚢内の精子数の合計は、交尾後15分で約8万、20分で約10万、以後40-90分の間は変化がなく、90分で増加し始め、2時間で約14万に達すると報告した。この2時間目の数は、未交尾雄蛾の貯精嚢および輸精管膨大部(原著者はこれをそれぞれ射精嚢および貯精嚢と呼んでいる)内における精子数(約104万)の約13.5%に当ると云う。
射精は60-90分の間隔で行なわれ(498,863)、射精には、上述のように、毎回約40分かかるから、第1回目の交尾(初交)は1時間で足り、引き続きその雄蛾を再交させる場合には2時間交尾させておけばよいことになる。
第1回と第2回との射精における精子の量が等しいか否かは測定がむずかしいが、大村は射出された精莢を秤量して(第11表)、その値が貯精嚢に流入する精液量の近似値を示すものと考えている。
| 試験 | 交 尾 | 前回の交尾 との間隔 |
調査 個体数 |
精莢の重量 | 比 | 雄蛾の状態 |
| 1 2 3 4 5 6 |
初 交 再 交 〃 〃 三 交 初 交 |
-時間 5 10 24 24 - |
30 8 8 8 6 9 |
0.0085g 0.0031 0.0030 0.0049 0.0036 0.0132 |
100 36 35 58 42 |
化蛾後約20時間 試験1に用いたものの一部 〃 〃 試験1および2に用いたもの 約15℃で化蛾後約12日 |
町田・渡辺(498)によれば、再交の射精継続時間は初交より短く、初交では終了までに30分内外かかるが、再交においては17.6分(6例平均)であった。
生理食塩水中に取出した精子の生存力からみると、貯精嚢および輸精管膨大部からとったままの非活動性の精子は、これに射精管下部の分泌液を加えて活動性にしたものに比べて冷蔵に耐え、化蛾後5℃に10日間冷蔵した蛾からの精子の生存力は化蛾当日のものからとった精子と異ならないが、20日間冷蔵した蛾の精子は生存力が劣ったと云う(751)。
精子の生理学的研究には見波(532)の報告もある。
b 外部生殖器官
雄蛾の外部生殖器官を第13図に示す。

第13図 雄蛾の外部生殖器官(模式)
A:第8腹節背板、B:第10腹節背板、C:補握器、D:竜骨
E:繋帯、F:陰茎、G:第8腹節腹板、H:肛門、I:鈎器
捕握器(攫握器)は交尾の際に雌蛾の第8腹節腹板の鋸歯板内側に引かけて連結を確実にする器官である。鈎器と竜骨も同腹板後縁中央の彎入部を挟んで連結の役目をする。
雄蛾の外部生殖器官の原基については竹田・田中(981)の研究があり、幼虫の腹部第8節腹面の後方約3/4と第9節の一部に亘る、腹線を中心とした部分の皮膚から陰茎と繋帯、左右の上腹線を中心とし基線を含む部分の皮膚から左右の捕握器、左右の尾脚から鈎器がそれぞれ形成されると云う。
C 雄生殖器官の異常
交配の実際問題にも関係して興味のあるのは梅谷・大村(1117)の異常精莢である。遺伝的な異常であるが、この系統の特異な点は、雄の初交において不受精卵が非常に多く、再交では受精率が高まると云う常識に反する結果を示すことである。射精の際に、射精管上部からの分泌液が精莢を形成し、精子はこれに包まれて交尾嚢内に射出されることを既に述べたが、正常の場合には、この精莢の頚部後端の開口が精子管の入口に近接していて(第14図)、精子が精莢から出て精子管に入り易いようになっている。
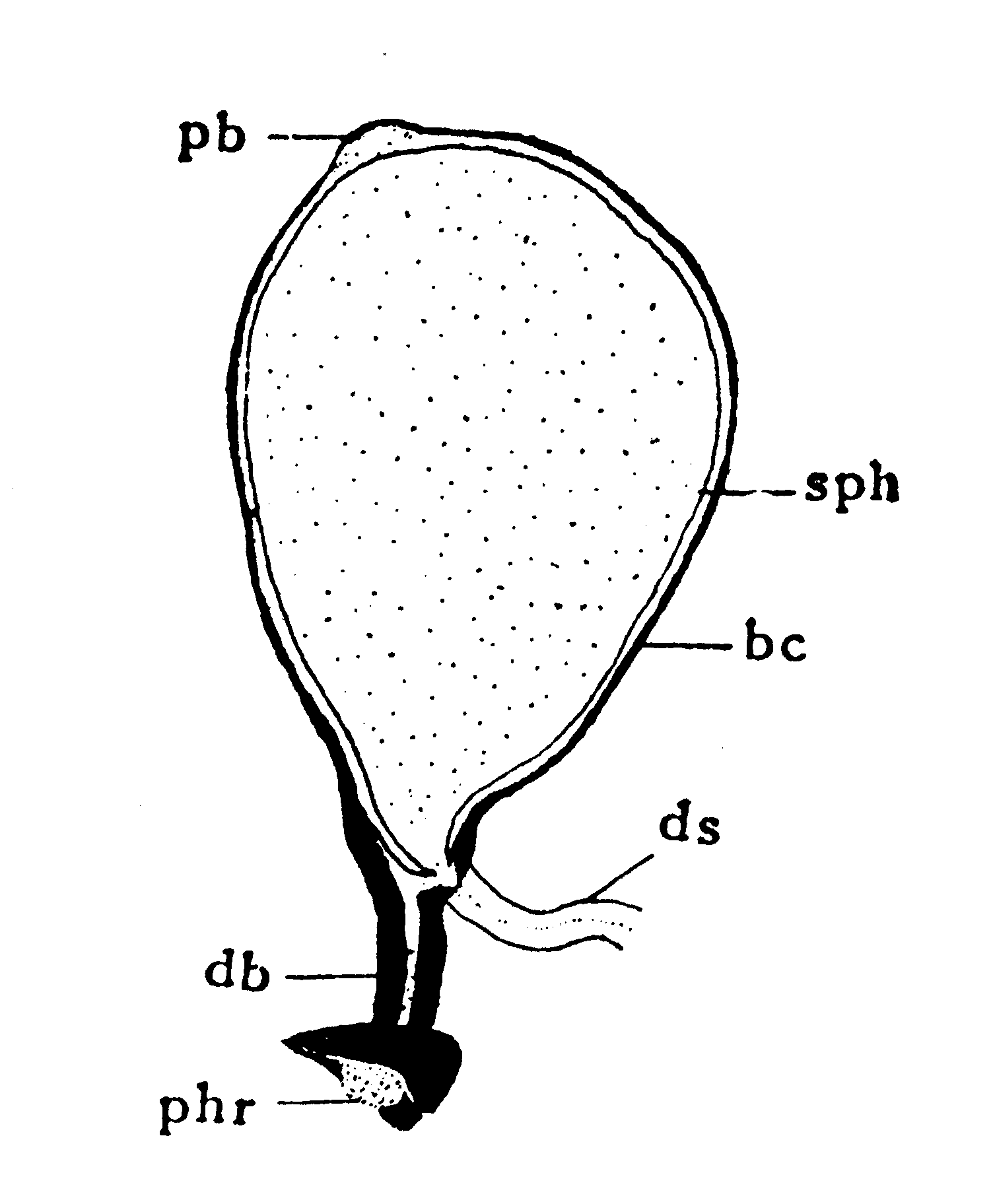
第14図 正常な精莢(模式)(Omura(760))
bc:交尾嚢、db:交尾嚢導管、ds:精子管、pb:真珠体(射精管中部からの分泌物)
phr:交尾孔閉塞栓、sph:精莢
処が、この異常精莢系統の雄蛾の初交の場合には、精莢の頚部が非常に長く、交尾嚢導管の中に入り込み、甚しい場合には交尾孔の外にまで出ており(第15図A)、精子が精子管に移りにくい状態になっている。
同じ雌蛾に同じ異常雄蛾を統けて再交すると、第2回目の交尾による精莢は初交のものに比べて柔軟で、頚部が内方へ押込められ、その開ロ部が精子管の入口に近い位置に来ているものが多かった(第15図B)。再交の受精率が高いのはこのためであろう。初交の場合でも、長時間交尾させておくと受精率の高まるのは、自然に離れて再交尾するものができるためと考えられる。同じ雌に違う初交の雄を統けて交尾させても受精率は改善されないが、この場合には、解剖してみると、2個の精莢の頚部が共に交尾嚢導管の中に入っていた。再交の精莢が柔軟になるのは前節(Ⅰ3Ba)で述べたことによって理解される。
第15図 異常精莢(模式)(梅谷・大村(1117))
b:交尾嚢、d:交尾嚢導管、s:精子管
A:同じ雌に同じ雄を2回続けて交尾させた場合の第1回目の精莢
B:同第2回目の精莢
鈴木・大村(890)は、産卵数が少なく受精率の非常に低い系統を調べ、雄蛾に原囚があって、精子が少なく、異常に縮れたものの多いことを認めた。この“ちぢれ精子”は無核精子であるが受精嚢にまで達している(Ⅰ3Ab)。
陰茎付属筋肉の退化によって不受精卵蛾の生ずる系統もある(1084,1122)。この異常の発現は蛹期の保護温度によって著しく影響される(Ⅲ5A)。
輸精管に突起または分枝の生ずる場合の多いことが報告されているが(96,760)、機能に障害があるか否かは記載されていない。
体の片側で、第8節の睾丸に2精室しかなく、その代りに、第7節に1個余分の睾丸(1精室)のある畸形が報告されているが(267)、これは生殖腺の発生過程(Ⅰ1)を考えれば成因が理解される。体側に瘤状の突起があり、その中に遊離した睾丸の含まれている畸形(493,494)は卵巣の畸形として記載されているものと(Ⅰ2Cc)同じ性質の発生異常である。
異常生殖腺蚕(Ⅰ2Abi)は睾丸の精室数も多い。雌雄モザイクについては既に述べた(Ⅰ2Cb)。
原蚕飼育が普通養蚕とは原則的に違ったものであるかのような響きを持った種繭養蚕、糸繭養蚕と云う言葉が使われているが、木暮(407)も述べているように、この二つが原則的に違った飼育法によるものと考えるのは正しくない。また、分場と云う特別な世界の中で行なわれてきた原蚕飼育が、従来の方法による種繭養蚕では維持できなくなってきた現状においては、この二つを全然別なものと考えることは実情にも合わなくなってきているように思われる。
原蚕飼育ならびに蚕卵の増産については、古くから、多くの経験者によって数多くの見解や試験結果が発表されており、今更何を述べても云い古されたことの埒外に出ることは殆ど不可能である。しかしこれらを検討し、また簡潔明快な木暮の記述(407)、諸星(596)、 山口(1159)の綜説などを通じて感じられるのは、非常に多くの、時には全く相反する事実や解釈が示されていて、これらを貫く原則的なものが何であるかをつかみにくいことである。
ここでは、問題を造卵および産卵と云う一点に集約して、この点から原蚕飼育の原則的な検討をこころみたい。
1 体内環境と造卵および蚕卵
産卵数を直接に規定する条件は造卵数と産卵歩合とであるが、このうち、造卵数は初めに分化する生殖細胞数とも蛹期のなかば頃までの卵母細胞の数とも関係がなく、蛹期の中期頃から影響の現われる“ある必要な栄養物質”の量によってきまるらしいことを前章(Ⅰ2Abi)で明かにした。原蚕養育の影響は必らずこの体内環境を通して結果に現われるのであるから、先ず第一に体内環境を考えなければ、造卵数または産卵数を正しく理解することはできない。
A 蛹重
造卵数またぱ産卵数が母体に蓄積された栄養に関係があるとすると、当然、母体の大きさと造卵数との間に関係がある筈であるが、雌蛹の体重と産卵数との間には明かに正の相関があり(第12表)、普通には、蛹が大きいと産卵数が多い。蛹重は繭層重と相関があるので、繭層重と産卵数との間にも相関があるが(772)、蛹重と産卵数との相関よりも蛹重と産卵量との相関の方が強く(889)、数と量との問題(Ⅰ2Abi)の関係していることを示している。普通には、蛹が大きいと産卵量ばかりでなく、卵1粒の重量も大きくなる(572)。
| 蛹重量(g) 産卵数(粒) |
0.8 | 0.9 | 1.0 | 1.1 | 1.2 | 1.3 | 1.4 | 1.5 | 1.6 | 1.7 | 1.8 | 計 |
| 250 300 350 400 450 500 550 600 650 700 |
- - - - - - - - - - |
- - 1 2 - - - - - - |
- - - 1 2 - - - - - |
2 1 4 7 6 3 1 - - - |
1 1 1 6 17 13 2 2 2 - |
2 - - 5 7 11 22 4 2 - |
- - - 7 5 11 10 9 1 - |
- - - 1 1 5 9 11 3 2 |
- - 1 - 4 2 4 4 1 1 |
- - - - 2 1 - - - 1 |
- - - - - - - - 1 1 |
5 2 7 29 44 46 48 30 10 5 |
| 計 | - | 3 | 3 | 24 | 45 | 53 | 43 | 32 | 17 | 4 | 2 | 226 |
しかし、蛹重が増加しても産卵量(あるいは数)はこれと同じ割合では増加せず、後者の増加が幾分小さいので、雌蛹1kg当りの産卵量を比較すると,蛹重のおもいものは頭数が少ないため、蛹重の軽いものよりも却って産卵量の少ない傾向がある(14)。 従って、雌雄を区別しない種繭1kg当りの産卵量も、繭重の重い場合に多いとは限らない(第13、14表)。
この成績は、農林省蚕糸試験場の新庄、小淵沢、宮崎の3原蚕種製造所が協力して行なった5令期給桑量試験の結果であって、試験区によって繭に軽重があるが、この区間の平均繭重の変動が個体変異の場合(第12表)と同様に、1蛾当りの産卵量と正の相関を示している。しかし、この相関は同じ飼育場所内での区間に限られ、飼育場所が違えば、平均繭重は同じでも産卵量の全く相違する場合が少なくない。
| 蚕品種 | 飼育場所 | 5令給桑量 (kg) |
a 繭重 (g) |
b 種繭1kg当り 粗卵量(g) |
c ♀1蛾当り 粗卵量(g) |
繭層 歩合 (%) |
蛹重 (g) |
正常卵 1g粒数 |
c/a ×100 |
| 日124号 (春蚕) |
新庄 | 21 18 15 13 |
1.85 1.81 1.70 1.54 |
67 72 80 72 |
0.28 0.29 0.27 0.26 |
23.8 23.4 23.5 23.9 |
1.41 1.39 1.30 1.17 |
1,625 1,651 1,650 1,690 |
15 16 16 17 |
| 小淵沢 | 22 19 16 13 |
1.74 1.66 1.59 1.48 |
96 105 96 101 |
0.33 0.32 0.30 0.29 |
23.1 23.4 23.3 23.0 |
1.37 1.27 1.22 1.14 |
1,677 1,705 1,733 1,775 |
19 19 20 20 |
|
| 宮崎 | 23 20 17 15 |
1.98 1.95 1.88 1.76 |
95 99 103 108 |
0.34 0.34 0.35 0.34 |
22.7 22.7 23.0 23.0 |
1.53 1.51 1.45 1.36 |
1,632 1,635 1,652 1,679 |
17 18 19 20 |
|
| 支124号 (春蚕) |
新庄 | 20 18 14 12 |
1.84 1.75 1.55 1.52 |
55 59 69 55 |
0.27 0.25 0.26 0.23 |
21.7 21.8 21.8 21.7 |
1.44 1.37 1.21 1.19 |
1,645 1,700 1,735 1,735 |
14 14 17 15 |
| 小淵沢 | 22 18 16 14 |
1.46 1.42 1.34 1.28 |
74 65 68 74 |
0.24 0.22 0.21 0.21 |
22.0 21.7 21.7 21.9 |
1.14 1.11 1.05 1.00 |
1,813 1,800 1,833 1,830 |
17 15 16 16 |
|
| 宮崎 | 23 20 18 15 |
1.92 1.89 1.81 1.68 |
74 67 68 74 |
0.28 0.28 0.28 0.25 |
21.4 21.2 21.5 21.9 |
1.53 1.51 1.45 1.36 |
1,599 1,608 1,627 1,667 |
15 15 16 15 |
| 蚕品種 | 飼育場所 | 5令給桑量 (kg) |
a 繭重 (g) |
b 種繭1kg当り 粗卵量(g) |
c ♀1蛾当り 粗卵量(g) |
繭層 歩合 (%) |
蛹重 (g) |
正常卵 1g粒数 |
c/a ×100 |
| 2・4 (秋蚕) |
新庄 | 21 18 15 12 |
1.83 1.76 1.63 1.39 |
82 80 82 74 |
0.32 0.31 0.29 0.25 |
22.0 21.6 21.5 20.9 |
1.43 1.38 1.28 1.10 |
1,590 1,620 1,645 1,670 |
18 18 18 18 |
| 小淵沢 | 21 18 15 13 |
1.75 1.71 1.58 1.52 |
101 109 100 101 |
0.37 0.35 0.33 0.32 |
22.5 22.4 22.7 22.5 |
1.36 1.33 1.22 1.18 |
1,623 1,660 1,666 1,670 |
21 20 21 21 |
|
| 宮崎 | 23 20 18 15 |
1.72 1.66 1.59 1.50 |
96 100 100 89 |
0.31 0.30 0.28 0.25 |
21.6 21.7 21.9 22.0 |
1.35 1.30 1.24 1.17 |
1,721 1,713 1,750 1,756 |
18 18 18 17 |
|
| 5・4 (秋蚕) |
新庄 | 21 18 15 12 |
1.76 1.70 1.54 1.39 |
93 93 87 85 |
0.34 0.34 0.33 0.26 |
20.2 20.0 19.9 19.5 |
1.41 1.36 1.23 1.12 |
1,630 1,645 1,670 1,705 |
19 20 22 19 |
| 小淵沢 | 20 18 16 13 |
1.69 1.71 1.62 1.57 |
100 92 84 100 |
0.37 0.32 0.36 0.35 |
19.3 19.3 19.4 19.3 |
1.36 1.38 1.31 1.27 |
1,624 1,626 1,633 1,644 |
22 19 22 22 |
|
| 宮崎 | 23 20 18 15 |
1.53 1.54 1.52 1.47 |
88 81 92 85 |
0.29 0.28 0.27 0.26 |
19.7 19.5 19.4 19.5 |
1.23 1.24 1.23 1.18 |
1,712 1,712 1,697 1,710 |
19 18 18 18 |
第13表の日124号および支124号においては、繭重の増加に連れて繭重1g当りの産卵量(c/a)が減少する傾向を示しているが、第14表(交雑原種)においてこの傾向の認められないことは興味がある。これが原種と交雑種との相違を示すものか否かはなお明かでないが、竹内ら(996)の支122号×日122号および支115号×日122号についての成績から計算してみても、繭重1g当り産卵数は繭重が増加しても減少してはいない(Ⅱ2Ab)。
渡部(1133)は雌蛹の体重1g当り産卵数によって産卵効率を表わし、原種に比べて交雑種の効率の高いことを示したが(第15表)、上記の現象は、この効率が原種では一定でなく、蛹重(または繭重)の増加に連れて低下するが、交雑種においてはこのような効率低下の起こらないことを意味するものかも知れない。これに対して、繭層歩合は繭重の増減に拘らず殆ど一定であるから(第13、14表)、原種においては、卵の造成に関する代謝効率と絹物質造成に関する代謝効率との間に相違のあることがうかがわれる。
正常卵1g粒数は蛹重または繭重の軽くなるに連れて多くなるが、この傾向には原種と交雑種との間に特別な違いが認められないようである。
| 蚕品種 | 調査蛾数 | 産卵効率 | 変異係数(%) |
| 日1号 支4号 日122号 カンボージュ |
43 45 44 65 |
325 295 324 441 |
13 33 21 29 |
| 日1号×支4号 支4号×日1号 支4号×日122号 日122号×支4号 日1号×カンボージュ 支4号×カンボージュ |
29 73 93 90 84 83 |
392 366 366 385 439 448 |
7 10 10 8 13 10 |
B 卵の造成と絹物質の造成との関係
浜崎(149)は、3令末期または4令初期に卵巣を摘出した蚕は初めは体も小さく、斃蚕が多いが、5令2日目頃から回復し、残ったものの繭層重および蛹重は対照区より大きかったと報告している(第16表)。永井(631a)は、これについて、幼虫の卵巣は体重に対して極めて小さな割合を占めているに過ぎないにも拘らず、その摘出が蛹重に影響するものならば、変態ホルモンとの関連があるのではないかと考えている。藤本(101)は、栄養状態が正常の場合には卵巣の存否は絹糸腺の発育に影響をおよぼさないが、絶食させて栄養不足の状態にすると卵巣の減少よりも繭層重の減少が大きいことからみて、卵巣の存否および発育は絹糸腺に影響をおよぼすものであろうと考えている。
| 昭 和 | 新白×日110号 | ||||
| 対 照 区 | 去 勢 区 | 対 照 区 | 去 勢 区 | ||
| 繭層重 | ♂ ♀ |
111mg 94 |
124mg 102 |
173mg 170 |
197mg 186 |
| 蛹 重 | ♂ ♀ |
841 955 |
889 981 |
987 1,212 |
1,022 1,233 |
小原(388)によれば、蚕の体液蛋白の電気泳動像は5令の中頃から雌雄差が明かになるが、5令期に卵巣を摘出した蛹について泳動像を調べたところ、一見、雄に似た像が得られたと云う。
これらの結果は、卵の造成と絹物質の造成との聞に何らかの関連のあることだけは示しているが、直接的な物質の動きとしては明かでない。
これに対し、絹物質造成のための素材を卵の造成に転用することができるか否かについては、赤尾(33,34)の実験結果がかなり具体的な解答を与えている。
4令期に絹糸腺を摘出しておくと、蚕は節聞がふくれて膿蚕様の形態を示し、大部分が化蛹前に死んでしまう。これは絹蛋白を合成する材料であるアミノ酸が、吐糸によるはけ場がないために、体液中に蓄積してアミノ酸過剰症を起こすためであるが、同時に、吐糸によって大量に排出される筈の水分も体内に残留して生理障害の原因になっている。蚕にとっては、絹糸腺は云わば過剰なアミノ酸、水分、カルシウムなどの排泄器官として働いている訳であるから、これを除去すると排泄不能のための生理障害が起こるのである。
処が、この除腺蚕を緑蚕上蔟させると(第17表)、これらの障害が軽減し、緑蚕上蔟の早いものでは、同時に上蔟させた除腺しない緑蚕上蔟に比べて化蛹歩合および化蛾歩合が高く、産卵も多い。これは、本来ならば絹の生成に向けられる筈の過剰の蓄積物が発育、造卵および体を維持するためのエネルギー源に転用されたために、過剰蓄積を持たない非除腺蚕よりも生理上有利な立場にあったことを示すものと考えられる。この場合、除腺蚕のうちでも、過剰蓄積物を卵の生産のために消費し得る雌の方が化蛹歩合、化蛾歩合ともに高いことは興味がある。
| 緑蚕上蔟 の時期 |
処理 | 性別 | 熟蚕頭数に対 する化蛹頭数 |
化蛹頭数に対 する化蛾頭数 |
産卵数 (平 均) |
残留卵数 (平 均) |
| 5 令 4日目 |
無 処 理 | ♀ ♂ |
29% 30 |
71% 75 |
41粒 - |
10粒 - |
| 吐糸孔閉鎖 | ♀ ♂ |
28 28 |
28 0 |
141 - |
1 - |
|
| 除 腺 | ♀ ♂ |
78 68 |
77 82 |
264 - |
2 - |
|
| 5 令 5日目 |
無 処 理 | ♀ ♂ |
53 80 |
57 80 |
287 - |
17 - |
| 吐糸孔閉鎖 | ♀ ♂ |
28 33 |
0 0 |
- - |
- - |
|
| 除 腺 | ♀ ♂ |
95 81 |
95 88 |
459 - |
5 - |
|
| 5 令 6日目 |
無 処 理 | ♀ ♂ |
81 88 |
90 95 |
403 - |
55 - |
| 吐糸孔閉鎖 | ♀ ♂ |
35 8 |
0 0 |
- - |
- - |
|
| 除 腺 | ♀ ♂ |
85 50 |
73 66 |
551 - |
32 - |
|
| 5 令 7日目 |
無 処 理 | ♀ ♂ |
80 91 |
95 77 |
551 - |
36 - |
| 吐糸孔閉鎖 | ♀ ♂ |
8 4 |
0 0 |
- - |
- - |
|
| 除 腺 | ♀ ♂ |
33 0 |
50 0 |
352 - |
207 - |
|
| 5 令 8日目 |
無 処 理 | ♀ ♂ |
84 96 |
90 91 |
626 - |
94 - |
| 吐糸孔閉鎖 | ♀ ♂ |
0 0 |
0 0 |
- - |
- - |
|
| 除 腺 | ♀ ♂ |
4 4 |
0 0 |
- - |
- - |
このように、絹物質造成のための素材も場合によっては卵の造成に転用できるものと考えられるが、正常蚕においても同様な転用が行なわれ得るか否かは、蚕種製造上大きな関心の寄せられる問題である。
平板吐糸蚕は正常吐糸蚕の70-80%ぐらいしか吐糸せず、化蛹の頃に解剖してみると、絹糸腺内に多量の絹物質の残留しているのが肉眼的にも認められるが、やがてこれが分解して腺外に出るため、体液中のアミノ酸が増加し、化蛹後3-4目で最高に達する。この頃は、丁度、卵巣が急激な発育を始める時期に当るから(Ⅰ2A
bi)、もし体液中に移行した絹糸腺内容の分解物が卵の造成に転用され得るならば造卵数が増加する筈であるが、体内に含有されているグリコーゲンの消費の減少することからみて、絹物質分解物がエネルギー源として一部用いられるのであろうと考えられる以外は、大部分尿酸に変って直腸内に蓄積されるもののようである。また一部が有毒なアミンに変って卵細胞の発育を妨げている可能性もあり、平板吐糸蚕の造卵数は正常吐糸蚕よりも少ない(32,35,36,711)(第18表)。
| 試験区 | 個体番号 | 造 卵 数 | ||
| 左側卵巣 | 右側卵巣 | 計 | ||
| 標準区 | 1 2 3 4 5 |
300 294 319 323 337 |
300 303 303 321 305 |
600 597 622 644 642 |
| 平板吐糸区 | 1 2 3 4 5 |
212 276 237 242 240 |
217 274 237 251 260 |
429 550 474 493 500 |
吐糸孔閉鎖蚕の成績が、緑蚕上蔟を行なっても除腺蚕より悪い(第17表)理由は明かでないが、絹糸腺の有無によって転用率に相違があるものとすれば興味がある。大宮(739)も、上蔟後2日目に吐糸口を閉鎖したものは、3日目または4日目に閉鎖したものよりも造卵数の少ないことをみている。
多糸量系品種ほど、正常吐糸の場合にも残留絹が多く、卵巣、卵細胞の発育が悪いと云われていることは(631)、上に述べた実験結果と関連のある問題と考えられる。
絹物質の造成と卵の造成との間に素材の転用が普通に行なわれるものならば、両者の聞には競合も起こり得る筈であるが、繭層重と産卵数との聞に正の相関のあることは(Ⅱ1A)、正常の場合には両者の間に競合関係の少ないことを示すものである。
種繭養蚕は繭を目的とするものではないが、以上の結果から考えて、造卵数を多くするためには、蚕が十分に吐糸できるような条件を整える必要のあることがわかる。
種々な発育時期の蚕に放射性同位元素(41C)で標識した桑を与え、その蚕の吐いた繭糸を調べると、仮りに繭糸の長さが1,200mあるものとすれば吐き初めから100mは5令4日目、200-400mは同6日目、700-1,000mは同8日目(熟蚕直前)および5令初・中期に食べた桑からの材料によって作られていると云うように、食べた時期によって、その桑が絹の造成のために使用される時期が異なり、また、4日目以降に食べた桑からの材料は直接に(直接的生成)、それ以前の分は、一旦体組織に取入れられたものが変態に伴って移行して(間接的生成)繭繊維を作ることがわかった(139a,140)。この結果から、繭繊維は主として5令の中・後期に食べた桑を材料にして作られるものと考えられる。卵の造成についても、同様な実験の結果から、5令の初・中期および4令期に食べた桑が主として材料に用いられる(140)と云われている。何時頃食べた桑が卵の造成に最も大きく影響するかは蚕種製造上関心の深い問題であるが、この実験結果の解釈には検討の余地がある。
繭糸の場合には、放射性桑を与える時期の相違によって、放射能の検出される繭糸の部分が異なるので問題はないが、卵の場合には、どの時期に放射性桑を与えても放射能は卵巣または卵全体に現われて、部分の区別がなく、検出される放射能の強さが、与える時期によって相違するだけである。
この実験においては、1頭の蚕に放射性桑1cm2(その放射活性は平均458,000c/m/cm2)ずつを食べさせたのであるが、卵に現われた放射能(対1蛾)の実数を第19表a側、これを5令3日目を100とする指数で表わしたものを同a’欄に示す。
| 放射性桑を 与えた時期 |
a | a’ | b | b’ | c |
| 卵の放射能 (対 1 蛾) |
同指数 | 消化量(g) (対♀100頭) |
同指数 | 寄与した 消化量指数 |
|
| 4令起蚕 4令中 5令起蚕 2日目 3日目 4日目 5日目 6日目 7日目 |
18,564c/m - 19,701 - 22,644 18,275 18,560 16,324 7,544 |
82 (87)* 87 (94)** 100 81 82 72 33 |
19.5*** 9.3 13.0 22.2 22.5 24.6 27.3 21.5 |
88 42 59 100 101 111 123 97 |
77┐ 31│ 56├345 100│ 81┘ 91┐ 89├212 32┘ |
この数字だけをみている限り、卵は主として5令の初・中期および4令に食べた桑を主な材料にして作られているように考えられるが、蚕は放射性桑1cm2のほかに普通の桑を十分に食べたのであるから、放射性桑はいわば普通の桑で稀釈された形で消化、吸収されたものと考えなければならない。摂取する桑の量は蚕の発育時期によって著しく相違する。この実験の場合の日々の食桑量は記載されていないので、品種は異なるが、日115号×支108号の消化量(517)をかりると、その実数は第19表b欄、5令3日目を100とする指数は同b’欄の通りである。消化量の多い時期には、それに比例して放射性物質の稀釈度が高く、これが卵巣に取込まれて同じ強さの放射能を現わすためには、稀釈度に比例した量の物質が取込まれる必要があるから、a’とb’とを用いて、造卵のために寄与したと考えられる消化量の指数を時期別に求めることができる。これをc欄に示した。この計算によれば、5令4日目までの寄与指数合計は345、5令5日以後の合計は212で、卵の造成のための材料は、5令5日目以後に食べた桑から、40%近いものがくることになる。
勿論、蚕の発育時期によって、食下した放射性桑の稀釈や組織への留存率、その濃縮などの条件が異なり、このような簡単な計算は適用できないかも知れないが、5令後期に食べた桑も卵の造成にかなり役立っていることは間違いないものと思われる。
なお、この実験の結果では、どの時期に放射性桑を与えても、放射能は雌蛾の卵管において基部から先端までの総べての卵細胞に均一に分布していたと云うから、退化卵の形成機構(Ⅰ2Abi)についてのこの面からの手がかりも得られなかった。
蚕の体内環境の問題は、別の言葉で云えばホルモンの問題、特に前胸腺ホルモンとアラク体ホルモンとの均衡の問題に帰着することが多く、これに食道下神経節からの休眠ホルモンの作用が加わって重要な体内環境を規定しているものと考えられるが(117,597)、 特別な項目としてこれを取上げず、必要に応じて言及する。
2 飼育環境と造卵および産卵
A 飼料
a 葉質
桑品種の相違が造卵または産卵に影響することは古くから云われているが、どのような桑品種がよいかと云うことについては必らずしも試験成績が一致せず、統一的な結論を出すことはむずかしい。しかし、これは、ただ卵の数(または量)にだけとらわれて、どうしてそのような結果になったかを考えないためである。結果を吟味してみると、殆ど共通的に、産卵の多い場合には蛹が健康で大きい。即ち、造卵のために特殊な効果があったと云うのではなく、蚕を大きく健康に育てた結果が卵の増産になったと云うのに過ぎないことが多い。
例えば、農林省蚕糸試験場新庄支場においては、蚕を赤木桑で飼うと産卵が多いと云われているが(675,739)、赤木で飼育した蚕は繭重が重くなっている(第20表)。産卵歩合には差がない。これは5月27日掃きの成績であるが、5月31日掃きも同様な傾向を示した。
| 蚕品種 | 桑品種 | 4令-結繭 減蚕歩合 |
繭 重 | 調査蛾数 | 造卵数 | 産卵歩合 |
| 日122号 | 赤木 剣持 |
6% 7 |
2.06g 1.93 |
54蛾 56 |
720粒 674 |
97% 97 |
| 支122号(太) | 赤木 剣持 |
1 1 |
2.04 1.79 |
61 44 |
718 681 |
99 99 |
剣持桑とこれをコルヒチンで処理して育成した4倍体桑との比較試験においては、4倍体桑で飼育したものの産卵数が多かったと云うが(838)、この場合にも4倍体区では蛹重、繭重が大きく、経過が短縮していた。
このように、蚕がよく育つ結果として産卵が多くなるのであれば、同じ桑品種を用いても、土地により時期によって成績の異なる場合がある訳で、桑品種の栄養価値を比較するつもりで行なった試験が、実際には桑葉の熟度試験や発育特性比較試験と同じことになってしまっていた場合があるかも知れないのである。
何か特殊な物質を桑に添加して与えると産卵が増加するのではないかと云う考えも古くからあるが、最近の試験で注目されるのは女王蜂の分泌する王乳(ローヤルゼリー)添食の成績である(173,205,700)。この結果(第21表)は、蚕が強健に育ち、産卵および繭層歩合は増加しているにも拘らず、蛹は特に大きくなっていない点で給桑量や桑品種による産卵増加の場合と相違している。王乳そのものの実用性は兎も角、産卵増加の研究の一環として、これらの点の究明が望まれる。
個々の蚕の産卵は増加しても、蚕が弱くなるようでは全体としての産卵増加は望めないが、上のように問題を分析すると、産卵増加と強健性とは相反するものではなく、蚕を強健に育てることが産卵増加の方法に外ならないことがわかる。 葉質問題に関して期待されるのは人工飼料による研究である。現在の処では、未だ、葉質の異なる桑葉粉末を加えて作った人工飼料で蚕を飼うと結果に差があった、と云うような常識的なことの確認の域をあまり出ていないが、これによって、結果の相違が桑成分の何処の違いによるもので、どうすればこれを改善することができるかと云う分析(47,281)をこれまでよりは科学的に進められるようになった。但し、普通に葉質と云う中には、栄養的な価値ばかりではなく、桑の萎れ易さと云うような物理的な諸性質までも含まれているのであるが、人工飼料による試験においては、これらの性質が均一になってしまうので、結果に現われる相違は、実際の場合よりも縮小あるいは拡大されている可能性がある。
β-カロチンおよびα-トコフェロールを人工飼料に添加すると産卵数が増加したと云う報告があるが(488)、現在の段階では、これらが特別に産卵増加に効果があったのか、これらの添加によって人工飼料が改善されたのかは明かでないように思われる。
経済的な問題や実施上の諸問題を技きにして、人工飼料によって蚕を飼うだけのことであれば、交雑種についてはかなり大規模な試験に成功しているが(510,883)、原蚕飼育も人工飼料によって十分行ない得ることが報告されている(282,306)。糸繭養蚕においては、凍霜害の場合の応急策としての人工飼料の利用がまず考えられるが、原蚕飼育においては冬季飼育への利用が考えられている。
農林省蚕糸試験場生理部(東京)において、桑葉粉末(50%添加)、馬鈴薯澱粉、脱脂大豆粕、セルロース粉末、寒天、ビタミンB群、ビタミンC、有機酸、防腐剤などを配合して作った人工飼料を用いて日124号および支124号を飼育、採種し、その次代蚕を同九州支場において桑葉を用いて飼育した結果は、九州支場において桑葉を用いて飼育、採種したものの成績と比べて、原種、交雑種共に殆ど差がなかった。
蚕卵数の増加した場合を調べると、蛹重の重い場合が多いことは前述の通りであるが、これから逆に、蛹重が大きければ産卵が多いときめることはできない。必要なのは蛹重そのものではなく、卵の造成に必要な物質の蓄積量であって、健康に発育した蚕においては蛹重の増加とこの蓄積の増加との間に相関があると云うことに過ぎない。
永井(627)は、蛹の充実度を蛹体の容積重、重量/容積×100,で表わし、その値の大きいものは産卵成績がよいと云っている。残絹や水分の残留のための蛹の肥大や、化蛹歩合や健蛹歩合を低下させるような蛹の肥大が卵の増産に結び付かないことは平板吐糸の場合(Ⅱ1B)をみても明かである。
原蚕には硬葉を与えて緊め飼いにするのがよく、硬葉を与えたものは軟葉を与えたものに比べて、繭重はほぼ同大でも採卵量が多い(307)と云われているのは、充実した桑で蚕を健康に育てるのがよいと云うのであって、一部で唱えられたように、過度の緊め飼いを行なって蛹を小さく作るのがよいと云うことではない。軟葉の害については、栄養の問題を離れても、三眠蚕(808)や、不吐糸蚕(660)、半化蛹蚕(827)の発生が多いなど蚕種製造上好ましくない結果が数多く報告されているから、軟葉給与を避けることは望ましいが、これが直ちに極端な緊め飼いがよいと云う結論に結び付くものではない。蛹が小さいと1頭当りの産卵量がへり、卵が小さくなることは既に述べたが、卵が小さいと、後に述べるように、浸酸その他の蚕種の取扱い上に難点が生じ、また次代蚕の成績にも影響する。
ただ、蚕には体内水分の調節作用が乏しいので(245,795)、多雨多湿な環境では、水分の乏しい桑を与えると体水分の平衡を回復するのに有効な場合もあり、また種繭の単位重量に対する採卵量は繭の小さい方が多いので(Ⅱ1A)、種繭を目方で購入する場合には極端な緊め飼いの有利な場合もある。
産卵の問題ではないが、きょう蛆卵の付着した桑葉は、高度晒粉の400倍液に30分間浸漬すると卵が離脱するから、引上げて余分の水滴を落し、そのまま給与すればよいと云う報告がある(1185)。
b 給桑量
両原種の体の大きさが、例えば欧18号と大造とのように著しく相違している場合を除けば、原種は交雑種に比べて桑の食べ方が少ないが、食べた桑の消化率は交雑種に劣らない(第22表)。
| 蚕 品 種 | 性別 | 食下量(g) | 消化量(g) | 食下率 (%) |
消化率 (%) |
||
| 生物量 | 乾物量 | 生物量 | 乾物量 | ||||
| 日122号 | ♀ ♂ |
14,935 12,627 |
3,627 3,070 |
5,876 4,918 |
1,427 1,192 |
51 46 |
39 38 |
| 支122号 | ♀ ♂ |
14,909 13,480 |
3,640 3,290 |
5,737 5,171 |
1,400 1,262 |
59 56 |
38 38 |
| 支115号 | ♀ ♂ |
13,630 12,422 |
3,327 3,032 |
5,161 4,435 |
1,260 1,082 |
55 52 |
37 35 |
| 日122号×支122号 | ♀ ♂ |
18,242 15,981 |
4,460 3,907 |
6,760 5,916 |
1,652 1,446 |
62 57 |
37 37 |
| 日122号×支115号 | ♀ ♂ |
18,085 15,510 |
4,422 3,786 |
6,672 5,780 |
1,631 1,411 |
62 58 |
36 37 |
即ち、良べた桑を消化する段階では、原種の効率は交雑種に劣らないが、食下量または消化量に対する卵の生産を比較すると先きに蛹重と生産卵重との関係について認めたと同様に(Ⅱ1A)、原種の効率が劣っている(第23表)。
| 蚕 品 種 | 産卵量(g) | 産卵乾物量歩合(%) | 産卵生物量歩合(%)* | |||
| 乾物量 | 生物量* | 対食下乾物量 | 対消化乾物量 | 対食下生物量 | 対消化生物量 | |
| 日122号 | 101.13 | 281 | 2.7 | 7.0 | 1.8 | 4.7 |
| 支122号 | 122.85 | 342 | 3.3 | 8.7 | 2.2 | 5.9 |
| 支115号 | 99.10 | 275 | 2.9 | 7.8 | 2.0 | 5.3 |
| 日122号×支122号 | 156.90 | 436 | 3.5 | 9.4 | 2.3 | 6.4 |
| 日122号×支115号 | 154.40 | 429 | 3.4 | 9.4 | 2.3 | 6.4 |
これから、卵1gの生産に必要な5令期の食下量および消化量を計算すると、蚕体の造成および維持に要する量をも含めて第24表のようになる。但し、これは雌だけを飽食させて飼育した場合である。給桑量を少なくすると食下率は上がるが、食下量、消化量、および消化率はともに低下する。原種についてのこの点の委しい調査はないが、交雑種についての試験結果は第25表の通りである。
| 生 葉 給桑量 |
食 下 乾物量 |
消 化 乾物量 |
繭 生産量 |
正 常1) 卵 数 |
正 常2) 卵 量 |
消化乾物量1g当り3) | ||
| 繭生産量 | 正常卵数 | 正常卵量 | ||||||
| 2,120g 92 84 76 68 60 |
391.0g 98 95 89 81 74 |
150.0g 93 90 84 73 66 |
211.0g 98 95 91 85 77 |
40,080粒 98 93 93 87 78 |
22.73g 97 92 84 73 66 |
1.41g 105 106 108 116 117 |
267.20粒 105 102 111 119 118 |
0.15g 104 102 100 100 100 |
これによってみると、対照区に比べて、給桑減量区の生産の絶対量は、繭重、卵数、卵量および繭層重(表には示してない)の何れも少ないが、消化量または給桑量、食下量に対するこれらの生産量の割合は、蚕の健康の低下しない範囲内では、減量区が対照区に劣らないか、またはこれよりも大きいことがわかる。
消化量が変化してもその1g当り正常卵量には殆ど変化のないことは、交雑種においては、繭重が増加しても繭重19当り産卵量の滅少しないこと(Ⅱ1A)を物質の動きの面からも確かめたことになる。
従って、蛹重または繭重と蚕卵量との関係は、給桑量-食下量-消化量-蛹重-産卵量を連ねる物質の動きの現れであることが理解され、桑を十分に食べさせて蛹を大きく作ることが産卵量増加の正道であることがわかる。
第13、14表に示した試験結果を給桑量の立場から考えると、表には示さなかった調査項目をも含めて、第26表のように概括することができる。
| 目的形質 | 最少限界給桑量 |
| 5令経過 減蚕歩合、健蛹歩合 繭 重 種繭収量 発蛾歩合 正常産卵蛾歩合 産卵量 対1蛾 〃 対種繭1kg 給桑量1kg当り精選卵量 |
18 kg 12-15 18 18 12-15 12-16 18 12-15 13-15 |
以上の試験における給桑量の減量は、毎回の給桑量を減らして1日4回(まれに3回)給与し、5令期間を通じての総給桑量を少なくしたのであるが、初め普通に給桑し、終りを打切った緑蚕上蔟の形で減量すると、総給桑量としては前の場合と同じでも、諸形質におよぼす減量の影響は遙かに大きい(第27表)。
| 区 別 | 給桑量 (kg) |
上 蔟 | 全繭重 (g) |
繭層重 (cg) |
正常卵数 (粒) |
造卵数 (粒) |
正常卵1万粒 重量 (g) |
| 毎回減量 給 桑 |
26(100) 21(81) 14(54) |
適 熟 〃 〃 |
1.75(100) 1.64( 94) 1.36( 78) |
38.1(100) 36.1( 95) 29.3( 77) |
599(100) 576( 96) 454( 75) |
613(100) 593( 97) 465( 76) |
6.119(100) 6.026( 99) 5.969( 98) |
| 緑蚕上蔟 | 26(100) 22(86) 17(66) 12(48) |
8日目適熟上蔟 7日目上蔟 6日目 〃 5日目 〃 |
1.75(100) 1.63( 93) 1.38( 79) 1.06( 61) |
38.1(100) 33.9( 89) 24.1( 63) 15.2( 40) |
599(100) 541( 90) 445( 74) 292( 49) |
613(100) 556( 91) 455( 749 297( 48) |
6.119(100) 6.189(101) 5.737( 94) 5.555( 91) |
第27表で注目されるのは、毎回減量区においては繭重、繭層重および造卵数がほぼ平行した減少を示しているのに対し、緑蚕区においては、繭重に比べて繭層重および造、産卵数の減少の著しいことである。この場合、繭層重の減少の最も著しいことは、繭繊維が主として5令中・後期に食べた桑を材料にして作られると云うこと(Ⅱ1B)から理解できるが、造卵数もまた毎回減量区よりも緑蚕区において減少の著しいことは、卵が主として5令の初・中期および4令期に食べた桑から作られると云う考えへの疑問を更に深めるようこ思われる。しかし、材料の準備はできていても、体の発育、維持および卵を造成するために必要なエネルギーの不足のために、間接に造卵数の減少することや、緑蚕上蔟においては影響が急激で調節作用の働きにくいことなども関係しているものと思われる。
大宮(737)は、1日4回給桑とし 6時間の食桑で僅かに残桑のある程度の給桑量を標準とし、これに対し食桑時間4時間10分および3時間で桑を除去する減量区を設け、数種の原種を用いて比較試験を行なった。この時間の比は10:7:5であるが、減量区の経過がのびたため、結局、食下量の比は100:92:81になった。6時間区と3時間区との間には1蛾当りの産卵数に差があったが、6時間区と4時間10分区との間には差が認められなかった。これは、上に述べた試験の結果から考えて、減量の程度が小さかったためと思われる。
c 葉質および給桑量と次代蚕
原蚕飼育の葉質および給桑量については、それが次代の蚕作にまで影響すると云う説と影響しないと云う考えとがあるが、古くから行なわれているこの点に関する論議は、葉質あるいは結桑量が不適当であると卵にどのような違いができるのか、これから孵化した蚕の性質は出発点においてどのように違っているのか、などの重要な過程を技きにして、ただ親の代に与えた桑の質または量と次代蚕の蚕作とを直接に結び付けようとしている場合が多く、悪かったから悪いと云うような論議になりがちであった。例えば、次代に減蚕が多いと云う結果が出た場合に、以前にはそのまま容認されたかも知れないが、減蚕の大部分が伝染性の病気によることが明かになった現在では、どうして次代蚕が病気にかかり易くなるのかの説明がなければ納得のゆく試験結果とは云われない。問題の解決にはこれらの中間過程を明かにすることが必要であるが、現在わかっていることは極めて少ない。
飼料の質や量が不適当な場合に卵に現われる最も明かな影響は、その重量からも、わかるように、卵の小さいことである(第13、14、27表)。
卵が小さければ、その卵殼も薄く、機械的な原因による障害があるのではないかと考えて、日122号および支124号を用い、5令食桑時間の約75%を経過したとき(5令5-6日目)に緑蚕上蔟させたものと,熟蚕になってから上蔟させたものとの卵の圧し潰し試験を行なったが、前者が特に潰れ易いと云うことはなかった(876)。 これは、卵殻は薄くなっても、同時に卵も小さいためかも知れない。
葉質不良の場合および緑蚕上蔟の場合には卵が浸酸によって潰れ易いことが知られているが(Ⅲ1A、Ⅷ2Aa)、これは、一般に、卵が小さいために起こる被害と考えられている。卵殻の厚さも関係しているのではないかと考えられるが大きさと卵殻の厚さとを区別した試験は行なわれていない。
親の栄養状態によって、完成卵の数および大きさの相違することは明かであるが、完成卵の卵黄成分に質的な変化かあるか否かについては、具体的には何もわかっていない。
このように、親の代の不適当な栄養状態が次代におよぼす影響の第一歩として、現在確認されているのは卵の小さいことだけである。卵の小さいことが、量ばかりではなく、同時に質的な相違を伴なっているか否かは今後明かにすべき問題である。ここでは、卵の小さいことを通してどのような問題があるかを検討する。
卵は小さくても、小さいなりに十分な栄養を貯えた卵であれば、これから孵化した蟻蚕は、体は小さくても発育においては劣らない筈である。品種による卵の大小はこの場合である。橋本(178)は、雄核発生(Ⅵ2A)を利用して、赤蚕と云う品種(赤蟻であるために孵化したときから区別がつく)の精核を卵の大きい欧18号およひ卵の小さい銀近江支の卵内で発生させ、孵化した赤蟻の体重を比較した。その結果によると、同じ赤蚕の精核だけに由来した蚕でありながら、初めのうちは、欧18号の卵内で発生させたものは体重が大きく、銀近江支の卵内で発生させたものは小さかったが、令を重ねるに連れて赤蚕本来の体重に近付き、幼虫末期には差がなくなった。この場合の銀近江支の卵は品種的に小さいだけで、栄養の質は正常と考えられるが、量が少なければ矢張り虫が小さくなるのである。鈴木・藤島(887)の調べた品種的な蚕卵の大小と壮蚕体重との関係においても同様のことが認められる。
これに対し、山口・市川(1160)は、日115号、支17号、支114号および欧19号につき、1蛾区内の卵を1粒ずつ秤量または大きさの測定をして大、中、小に分け、これから孵化した蟻蚕を1頭または大小別の群に分けて飼育した結果、同1蛾区内の卵の大小はそれから孵化する幼虫の健否と平行し、一般に、小卵からの蚕は虚弱で、2、3令頃まてに斃死するものが多く、大卵からの蚕は強健で結繭歩合が高いが、繭質に関しては中卵区が優れ、大卵区と小卵区との間には差がなく共に中卵区に劣った、と報告している。大、中、小の区分をどのような基準で行なったのかは明かでないが、結果からみて、この場合の小卵はただ形が小さいばかりでなく、質的にも何らかの欠陥があるかのように考えられる成績である。
池江(242)は1餓の産卵を産卵順序によって初、中、晩の3区に分け、卵の大きさ、孵化歩合、乾熱および冷蔵に対する抵抗力、蟻蚕の重量および絶食生命時数などを比較したが、これらは何れも初期産卵において最も大きく、中期がこれに次ぎ、晩期の産卵において最も小さかった。卵の減耗量はこれと反対に小形の晩期産卵において最も大きかった。飼育成績は、一般に初期産卵(大卵)が最もよく、中期がこれに次ぎ、晩期産卵(小卵)は最も悪かったと云う。永井(628)も同様な結果を報告している。
室賀(613)は1餓の産卵につき(欧9号)、卵の長さおよび幅を測定し、その積によって大きさを表わし、蚕卵の大小と催青日数の長短との関係を調べた。卵の大きさは1.52-1.92mm2であったが、温度22.5℃、 湿度75%の催青条件の場合に、催青着手から点青までの時間は中位の大きさ(1.66mm2)の区が最も短かく(244.6±0.5時間)、これより大小ともに長かった。点青から孵化までの期間は、一般に、卵の大きいほと長く、中位の卵が特に短いと云うことはなかった。合計すると催青着手から孵化までの期間は、僅かながら卵の大きいものが長かった。
その他一般に、小形な卵の飼育成績の悪いことが知られているから(535)、親の代の飼育条件が不適当な場合に次代蚕に悪影響のあることはこの点から一応肯定される。
しかし、大沢(773)が、新星×竜角の1蛾区414粒、新屋×褐円斑2が区、492粒および499粒、の3蛾区全部の卵を1粒々々産付け順に区別して1頭育を行なった結果によれば、3蛾区合計で斃蚕4頭に過ぎず(不孵化卵17粒)、産付けの早晩と飼育経過の長短、繭の大小、糸長、糸量、繊度などとの間には一定の関係がなかったと云うから、卵の大小に基ずく飼育成績の差は本質的なものではなく、飼育管理によって縮めることのできるものと考えられる。
第13、14表に示した給桑減量試験の場合には、2昼夜間産卵させた卵を初冬期に洗落し、比重選を施したので、不良卵は一応除かれている訳であるが、対照区と減量区との間に精選卵量歩合に差がなかったから、この程度の減量給桑では、特に不良卵が多くなることはないようである。また、その越年種を自然温度に保護しておき(新庄)、翌年4月中旬に胚が丙Bに発育したときに5℃に移し、30、60、90および120日間冷蔵して、各減量給桑区間に、冷蔵に耐える期間の違いがあるか否かを調べたが、日124号×支124号、その反交、2・4×5・4およびその反交の何れにおいても、給桑量の少ない区の卵ほど(卵が小さいほど)冷蔵に耐える期間が短いと云うような傾向は認められなかった。催青日数にも差がなかった。
| 蚕 品 種 | 区別 | 正常卵1,000粒 重 量 指 数 |
蟻蚕100頭重量指数 | |
| 春 | 初 秋 | |||
| 日124号×支124号 | 1 2 3 4 |
100 99 99 96 |
- - - - |
100 101 99 98 |
| 支124号×日124号 | 1 2 3 4 |
100 97 95 95 |
- - - - |
100 98 98 96 |
| 2・4 × 5・4 | 1 2 3 4 |
100 98 97 95 |
100 98 98 96 |
100 96 97 93 |
| 5・4 × 2・4 | 1 2 3 4 |
100 99 98 96 |
100 96 96 94 |
100 99 97 94 |
しかし、孵化した蟻蚕の体重は給桑量の少ない(卵の小さい)区ほど軽く(第28表)、その絶食生命時数も対照区に比べて幾分短かかった(第29表)。
小形な卵は、催青卵の冷蔵抵抗力(716)も催青中の乾燥抵抗力(744)も弱いと云う。
給桑減量区の蟻蚕の絶食生命時数の短い理由は、蟻蚕が元来弱いのか、体が小さいために消耗が比較的に大きいのかは明かでないが、第29表によって蟻蚕の死に方をみると、特別に弱いものが混っていてそれが早く死んだと云うのはなく、対照区に比べて、全体的におよそ半日ぐらい早く死んでいる。
| 蚕期 | 蚕品種 | 区別 | 孵化後の経過日数 | 調査条件 | |||||||||||
| 1日 | 2日 | 3日 | 4日 | 5日 | 6日 | ||||||||||
| Ⅰ | Ⅱ | Ⅰ | Ⅱ | Ⅰ | Ⅱ | Ⅰ | Ⅱ | Ⅰ | Ⅱ | Ⅰ | Ⅱ | ||||
| 春 | 2・4×5・4 | 1 2 3 4 |
0% 1 1 0 |
0% 0 0 0 |
0% 1 1 0 |
0% 0 0 0 |
0% 2 1 0 |
1% 0 1 0 |
2% 20 15 17 |
3% 13 9 21 |
48% 93 80 97 |
47% 89 82 95 |
100% 100 100 100 |
100% 100 100 100 |
蟻量 50mg宛 25℃ 78% |
| 5・4×2・4 | 1 2 3 4 |
0 0 0 0 |
0 0 0 0 |
0 0 2 0 |
0 0 0 0 |
0 0 6 0 |
0 1 1 1 |
2 2 17 6 |
0 1 2 13 |
33 38 90 57 |
44 60 32 47 |
100 100 100 100 |
100 100 100 100 |
||
| 初秋 | 2・4×5・4 | 1 2 3 4 |
0 0 0 0 |
0 0 0 0 |
1 0 0 0 |
0 1 0 0 |
3 0 1 1 |
3 3 3 2 |
6 3 7 3 |
7 12 13 6 |
94 91 99 97 |
92 98 99 95 |
100 100 100 100 |
100 100 100 100 |
100頭宛 25.8℃ 87% |
| 5・4×2・4 | 1 2 3 4 |
0 0 0 0 |
0 0 0 0 |
0 0 0 0 |
0 1 0 0 |
0 0 0 0 |
0 2 0 0 |
1 3 1 0 |
3 6 1 2 |
28 75 79 67 |
46 80 83 81 |
100 100 100 100 |
100 100 100 100 |
||
| 日124号× 支124号 |
1 2 3 4 |
0 0 0 0 |
0 0 0 0 |
0 0 1 3 |
1 2 0 2 |
0 1 2 7 |
3 4 1 3 |
33 32 45 36 |
23 31 24 47 |
92 98 96 97 |
91 97 97 98 |
100 100 100 100 |
100 100 100 100 |
100頭宛 25.9℃ 88% |
|
| 支124号× 日124号 |
1 2 3 4 |
0 0 0 0 |
0 0 0 0 |
0 0 1 0 |
2 0 3 0 |
0 0 4 4 |
6 2 10 2 |
29 18 34 39 |
35 33 36 60 |
100 96 98 100 |
100 100 99 100 |
100 100 100 100 |
100 100 100 100 |
||
この程度の違いならば、試験場のように、孵化当日に掃立て、その後の試育に注意すれば蚕作に影響することはない。
実際に給桑減量区の蟻蚕を掃立てた結果によると、最初に認められた蟻蚕休重の相違は3令起蚕においては殆ど解消しており、糸繭としての成績は、減蚕歩合(1-3令、4令-結繭とも)、収繭量、上繭歩合、上繭1㍑粒数、繭重、繭層重、繭層歩合などの総べての調査項目について対照区に劣らず、種繭としても種繭蚕数歩合、発蛾歩合、正常産卵蛾歩合、1蛾平均産卵数、精選卵量、精選卵歩合、正常卵1g粒数など、蚕種製造上関心のある項目について、親の代の給桑減量の悪形響は認められなかった。
大宮(741)は前項で述べた食桑時間の制限による5令期の減量給桑を6世代に亘って繰返えし、7代目を普通に飼育して比較したが、各区間に一定傾向の差を認め得なかったと云う。
しかし、農家の掃立てる蟻蚕のように2夜包み、3夜包み、ときには4夜包みも混じる場合には、上記の程度の絶食生命時数の違いも減蚕や遅れ蚕の原因となり、蚕作に影響するおそれは十分に考えられる。
この適例は小沢(721,722)の報告である。5令起蚕から、1日の食桑時間が16時間、12時間および6時間になるように、時間で食桑を制限した原蚕を用い、16時間区♀×12時間区♂、6時間区♀×12時間区♂の交雑によって次代蚕を作った。発蛾当日に掃立てたものでは区間に差がなかったが、蟻蚕を25℃に48時間おいた後に掃立てたものでは、減食区の次代蚕の飼育経過がのび、稚蚕期の減蚕歩合が多く、1-4眠期の体重も軽かった。5令以後の成績には差がなかった。また、減食区と緑蚕上蔟区との次代蚕を比較すると、ウイルスを添食しない場合には、当日掃きでは差がなく、緑蚕上蔟区の蟻蚕を25℃に24時間おいてから掃立てたものの成績が劣っていただけであったが、蟻蚕に4角形中腸型ウイルスを添食すると、当日掃きにおいても、緑蚕上蔟区は感染抵抗力が弱かった。25℃に24時間おいてから掃立てると、各区とも高い感染率を示した。
このように、親の代の不適当な給桑条件が次代蚕に影響する根拠として、現在確実なのは卵の小さくなることだけで、その卵の本質的な違いは明かでないが、これが他の各種の条件と組合わさって、二次的、三次的に蚕作に影響する機会を作ることは確かである。親の栄養が次代蚕に影響することは、この意味では十分に根拠がある。ただ、卵の本質的な違いよりも、二次的、三次的な影響による部分が大きいため、蚕種の保護、取扱い、催青、飼育などの条件によって、その現われ方が相違する。従来まちまちな結果の出ているのはこのためと思われる。実際問題としては、親の栄養に関心を持つと共に、1蛾の産卵の中にも、卵の大小など、これと同じ影響をおよぼす素因を持った卵が常に混っていることを考え、蚕種の保護取扱いに注意する必要がある。種繭の保護温度も卵の大小を通して蟻蚕の絶食生命時数に影響する(Ⅲ3Bb)。
B 飼育環境
a 温湿度
原蚕の飼育温度に関する試験結果は数多く発表されており、その細目においては必らずしも一致した結果ではないが、造卵または産卵に限って云えば、蛹の大きさとか健康とかをはなれて、造卵または産卵だけを特別に多くする飼育温度のないことは明かである。
例えば、大櫛ら(733)が恒温恒湿器を用いて、稚蚕から蔟中および種繭保護を通して一定の温湿度に保護した場合の成績(第30表)をみても、大体において、繭重の重いものは卵が大きく、産卵数も多いのであって、生理的温度の範囲内においては、飼育温度は体を作ることを通して産卵量に影響していることがわかる(大櫛らには、飼育中だけの温湿度を変化させた成績もあるが、僅かに30頭ぐらいの蛾について調査しているので、これをとらなかった)。
| 蚕品種 | 飼育温度 | 飼育湿度 | 蛹 重* | 1蛾当り普通卵数 | 普通卵1,000粒重量 |
| 支101号 | 23℃ 25 27 29.5 |
75% 70 80 72 |
0.93g 0.97 1.21 1.18 |
419粒 462 565 516 |
0.471g 0.469 0.503 0.498 |
| 支4号 | 23℃ 25 27 29.5 |
75% 70 80 72 |
1.21 1.22 1.45 1.32 |
495 503 543 502 |
0.526 0.553 0.562 0.510 |
| 日1号 | 23℃ 25 27 29.5 |
75% 70 80 72 |
1.31 1.27 1.50 1.41 |
486 528 597 485 |
0.575 0.596 0.591 0.600 |
| 欧7号 | 23℃ 25 27 29.5 |
75% 70 80 72 |
1.59 1.51 1.75 1.65 |
533 463 496 486 |
0.731 0.710 0.736 0.727 |
飼育温度の影響が間接的なものであるとすると、たとえ温度は違っても、食べた桑を同じように消化し、体を作ることのできる条件がととのえば、卵の生産は違わない筈である。この点に関して興味のあるのは清水ら(861)の成績である(第31表)。これは屋外条桑育などのために壮蚕期の温度の下がるのを想定して、その場合の食下量、消化量を調べた試験である。原種を用いての試験でありながら産卵に関する調査を欠いているのは残念であるが、17.5℃で飼育しても消化量が等しければ、繭生産も繭層生産も25℃飼育に劣らないのをみると、それとの相関から考えて(Ⅱ1A)、卵の生産も劣らなかったものと思われる。
| 蚕期 | 蚕品種 | 飼育温度 | 給桑量 (乾物) |
食 下 乾物量 |
消 化 乾物量 |
消化率 | 繭重 | 繭層重 | 健繭 歩合 |
|
| 4令 | 5令 | |||||||||
| 春 | 日124号 | 25℃ 25 17.5 |
25℃ 17.5 17.5 |
532g 617 566 |
327g 323 299 |
117g 111 88 |
35% 34 29 |
1.63g 1.62 1.51 |
39.3cg 37.1 31.1 |
94% 83 56 |
| 支124号 | 25 25 17.5 |
25 17.5 17.5 |
403 569 476 |
253 312 290 |
92 115 95 |
36 36 32 |
1.68 1.68 1.44 |
34.9 40.9 28.7 |
94 95 95 |
|
| 夏 | 日124号 | 23-25 | 25 17.5 25⇔17.5* |
622 680 611 |
315 293 309 |
76 73 85 |
24 24 27 |
1.22 1.21 1.22 |
25.0 25.4 25.9 |
93 87 85 |
| 支124号 | 23-25 | 25 17.5 25⇔17.5* |
549 566 547 |
313 323 318 |
82 85 86 |
26 26 27 |
1.19 1.09 1.28 |
25.3 25.3 24.9 |
94 90 92 |
|
| 晩秋 | 日124号 | 23-25 | 25 17.5 |
680 757 |
327 337 |
113 114 |
34 34 |
1.46 1.59 |
31.0 33.5 |
89 72 |
| 支124号 | 23-25 | 25 17.5 |
581 663 |
303 293 |
97 98 |
32 33 |
1.35 1.48 |
28.2 29.3 |
83 75 |
|
| 大 造 | 23-25 | 25 17.5 |
406 458 |
172 150 |
51 39 |
29 26 |
0.91 0.90 |
11.9 10.0 |
98 83 |
|
温度と消化量との関係は蚕の品種によって相違し、5令期の最適湿度が20℃付近のものも、25-26℃付近のものもある(515,1036)。稚蚕期にはこれよりも最適温度が高い。
しかし、温度は蚕の生理全体に影響し、各種器官の機能の均衡をも変化させるので、消化量以外のことにも十分注意しなければならない。正常に脱皮を重ね、変態し、化蛾するためには前胸腺ホルモンとアラタ体ホルモンとの働きが均衡を保っている必要がある。四眠蚕品種に縷々発生する三眠蚕は、不越年性になり易い催青をし、軟葉を給与し(186,803-808)、飼育温度が高いと(409)発生し易いことが知られていたが、福田はこれを前胸腺ホルモンとアラタ体ホルモンとの均衡の変化によるものとし、また中部糸腺が糸状に縊れる不吐糸蚕は、三眠蚕の出易い条件で飼育し、1-2令を高温(28℃)に遭わせると発生するが、3令あるいは4令後期の高温になって一層多発するのはホルモンの均衡の破れるためであると説明している(115,130)。ただし、別の不吐糸蚕においては、1-3令の高温(28-30℃)、4-5令の低温(20-22℃)が発生を多くすると云われている(660,907,909)。日124号に多発することのある半蛹(半化蛹蚕)は稚蚕期高温(28℃)、壮蚕期低温(22℃)で軟葉を給与した場合に多いと云う(827)。
大宮(739)によれば、日124号の造卵数は椎蚕期(1-2令)22℃ 、 壮蚕期(4-5令)28℃
の場合と稚蚕期28℃、壮蚕期22℃の場合とを比較すると後者に多いが、第1日目の産卵がやや少なく、残留卵歩合がやや高いと云う。この後者の条件は同じ場所で調査した上記の半蛹多発環境と一致するが、化蛹したものだけについての調査であるか否かについては記載されていない。
木暮(407)は、虫質その他をも考慮し、原蚕の飼育温度は、稚蚕期25-26℃、壮蚕期23-25℃が標準で、品種によって、例えば支那種はやや高めに、欧州種はやや低目にするのがよいと云っている。
1蛾当りの産卵の多いことは望ましいが、原蚕飼育にとっては、蚕の強健に育つことがそれ以上に大切で、原蚕飼育の省力化が問題になってきた今日では、蚕を丈夫に育てることが最も確実な産卵増加の方法と考えられる。
飼育温度の次代蚕への影響についても、二次的な条件が複雑に関与していることは葉質および給桑量の場合と同様である。第30表の成績は原蚕飼育温度の次代蚕への影響をみるために行なわれた大正13年の試験結果であり、大正12年にも同様(飼育中の温湿度だけを変化させた)の試験が行なわれているが、次代蚕に対する一定傾向の影響はみられなかった。
飼育湿度が蚕の生理に影響をおよぼすことは明かであるが(514,516)、温度と相関的に働くことが多く、産卵におよぼす飼育湿度単独の影響についての研究は極めて少ない。木暮(407)は70%ぐらいの湿度が産卵に最もよいと云っている。
湿度が高いと次代蚕に悪影響があると云われているのも、当代の不良条件を通しての二次的な影響と考えられる。
b 光線その他
蚕は明暗によって食下量および消化量が幾分相違し(第32表)、明飼育において多い。従って、蛹重、繭重なども明飼育において重い(第33表)。
| 令 | 食下乾物量(g) | 消化乾物量(g) | ||
| 明 | 暗 | 明 | 暗 | |
| 1 2 3 4 5♀ 〃♂ |
4 22 97 559 4,674 3,921 |
3 19 88 519 4,324 3,763 |
2 11 38 205 1,938 1,610 |
1 9 34 195 1,802 1,547 |
| 明 暗 | 性 | 食下乾物量 (g) |
消化乾物量 (g) |
対100粒(生物量) | ||
| 蛹 重(g) | 繭 重(g) | 繭層重(g) | ||||
| 明 | ♀ ♂ |
467 392 |
193 161 |
228 169 |
279 217 |
49 46 |
| 暗 | ♀ ♂ |
432 376 |
180 154 |
217 160 |
265 204 |
46 43 |
宮川(564)も明区の蚕は行動範囲が広く、食桑継続時間が長く、食下量の多いことを報告している。
これらの試験においては産卵調査が行なわれていないが、木暮(407)が、1-3令を1日18時聞以上照明した区は明かに産卵数が多かったと云っているのは、この食下量、消下量の増加によって説明することができよう。
給桑量を減らすと三眼蚕の発生が少なくなると云われている(808)ことと明るいと食下量の増加することとを考え合わせると、三眠蚕が高温、明飼育で多発する(409)理由の一部が理解される。福田・酒井(131)は、前項で述べた中部糸腺の異常による不吐糸蚕の発生が明暗によって異なり、4令期を明にしたときに異常が最も多く、3令期の影響がこれに次ぐと報告した。
明飼育では暗飼育に比べて有核精子/無核精子の比率が大きい(有核精子の割合が多い)と云うのも(Ⅰ3Ab)、あるいは蚕の食下量が多く、栄養がよくなるためかも知れない。
特定な時期に飼育するとか、特定な桑品種または多肥料桑、無肥料桑を給与すると云うような飼育環境を数代繰返えして、その影響を調べたり(740)、生産地、生産時期の異なる同一品種の卵を集めて、原蚕飼育環境の次代蚕への影響を比較する(718)などの試験も行なわれているが、これらの“環境”が単純でない上に、淘汰、選択、保護法などの問題が関係しているので、結果の分析はむずかしい。例えば、蚕種の孵化歩合一つをとっても、春採りと晩秋採りを違った産地から集めて比較する場合には、余程注意しなければ、原蚕飼育環境の影響ではなく、産卵後の保護法の違いや活性化の違いなどの影響をみていることになるおそれがある。
C 条桑育および屋外育
原蚕の飼育形式は、地方により、季節により、また飼育者によって、従来のままの蚕箔育で通している所から、雌雄鑑別の後、雄だけを条桑で飼う、雌雄ともに条桑で飼う、あるいは完全な露天育を行なうなど極めて種々であり、分場の事情の変化によって、条桑育に踏み切らなければならないとは考えながら、なお不安を残している蚕種業者も少なくないが、この点についての試験成績は不安を解消させるまでに至っていない。
この聞の事情は、緑川ら(527)の述べている次ぎの見解によっても窺われる。
1)蚕児の発育が斉一を欠く場合が多く、経過も延長する; 2)減蚕歩合、下繭歩合が多く、繭重軽く、対1万頭上繭収量が少ない; 3)特に日本種は環境条件に鋭敏で、下繭蚕が多く、繭中斃蚕も多い; 4)発蛾、交尾、産卵歩合は変らないが、不良蚕卵蛾歩合が高く、対1蛾の造卵数、産卵数が少なく、不良卵が多い; 5)雄に原因する不良産卵蛾歩合および不良卵混産歩合が高いが、これは温度に原因があると思われる; 6)卵の冷蔵に耐える期聞が短縮するが、これは雌に原因する処が大きい、などである。
これらの項目の中には、条桑育そのものが直接の原因であるとは考えにくいものも合まれているが、このようなことが直接にせよ間接にせよ起こるのは、桑を条桑で給与する方法そのものにあるのか、それとも温度その他の環境条件によるものかをまず明かにする必要がある。
谷口(1039)は、豊光および新玉を用い、5令2日目から次ぎの3区に分けて飼育を行なった。
対照(普通蚕室、補温)区 飼育温度 最高25.5℃ 最低21.5℃ 平均23.2℃
屋内条桑(蚕室2階)区 〃 27.0
18.0 22.2℃
屋外条桑(簡易ビニールハウス)区 〃 34.0 11.0 19.0
この場合、屋外条桑区は5令の3、4、5日目に降雨があり、雨漏りがして蚕体が濡れるような状況であった。結果は次ぎの通りである。
5 令 経 過 対照区に比べて屋内区は約1日、屋外区は2-2.5日長かった。
蚕 体 重 各区間に犬差なし。 、ヽ
5令減蚕歩合 豊光: 対照区2.0%、屋内区2.8%、屋外区10.7%
新玉: 〃 2.0%、 〃 1.5%、 〃 5.4%
繭 重 条桑の両区が対照区に比べてやや軽かった。
繭 層 歩 合 各区間に差なし。
対1蛾造卵数および産卵数 条桑の両区がやや少なかった。
不受精卵歩合 各区間に差なし。
対繭重造卵指数 ┐
対繭重産卵指数 ├対照区と屋内区との間には差がなく、屋外区は10-15%低かった。
対繭重正常卵数指数┘
1蛾の産卵数はやや少なくても、繭重がやや軽ければ対繭重当りの産卵数は対照区に劣らない筈のように考えられるが(Ⅱ1A)、屋外区の産卵指数が10-15%も低かったと云うことは、発蛾、交尾、あるいは正常産卵蛾歩合などに問題があったのではないかと思われる。この点の記載はない。
松田・石井(509)が日124号および支124号を5令2日目から雌雄別に飼育した結果によれば、数字は示されていないが、屋外条桑育は屋内普通育に比べて次ぎのような成績であった。
経 過 日 数 日124号が2日、支124号が1日おくれ,た。
健 蛹 歩 合 日124号は雌雄共に劣り、支124号は雌雄共に差がなかった。
繭重、繭層歩合 日124号は雌雄共に軽く、支124号は雌雄共に重かった。
発蛾歩合 ┐
対1蛾産卵数 │
受精卵歩合 ├日支共に差がなかった。
孵化歩合 │
次代蚕の性状 ┘
十万(297)が、瑞光と銀白とについて、全令普通育と全令条桑育とを屋内で比較した結果は次ぎの通りである。
瑞光 銀白
経 過 日 数 差なし 差なし
作 柄 差なし 条桑がやや勝る
対1蛾産卵数 殆ど差なし 条桑が勝る
掃立蟻量1g当り産卵数 条桑が繭かに劣る 条桑がやや勝る
但し、普通育は全令1日5回給桑、条桑育は1日2回(1-4令蚕箔条桑育)給桑であった。
5令だけの屋内条桑育と屋外茶桑育との結果の比較は次ぎの通りである。
瑞 光 銀白
経 過 日 数 屋外が2日おくれる 屋外が1日半おくれる
作 柄 屋外がやや劣る 差殆どなし
対1蛾産卵数 屋外が僅かに少ない 屋外が僅かに少ない
掃立蟻量1g当り産卵数 差少ない 差少ない
中島(650)は日122号および支122号(太)について試験し、普通育と条桑育とにおける造卵数の差は、厚飼いによって繭重に差が生じたために条桑育の造卵数が少なくなったものと考えている。種繭1kg当りの産卵量には普通育との間に差がなかった。
加納ら(308)は、原蚕飼育を屋外育に切りかえるための基礎資料を得る目的で、壮蚕の飼育温度28、23、18℃および屋外育(ビニールハウス、周囲を被覆せず)の比較試験を行なったが、28℃および18℃での飼育には減蚕が多く、健蛹歩合および産卵蛾歩合が低く、屋外育の方がこれらよりも良かった。また品種による差異も顕著であった。
これらの結果を通覧すると、最初に挙げた条桑育の問題点の多くは、条桑育に本質的なものと云うよりも気象その他の環境条件に基ずくことがわかる。従って、条桑育を取り入れる場合に大切なことは、環境整備と条桑育の取り入れとが経営の中でどのように調合されるかと云う点の検討にあると思われる。屋内条桑育ならば心配はない、と云う成績をあげている飼育者もある。
しかし、一面には、中島ら(651)が、日124号および支122号(太)の5令条桑育を23-24℃で行なった処、原因の不明な区間の成績変動や発蛾歩合の低下が認められたと云うから、ただ結果が良かったとか悪かったとかの比較だけではなく、問題点の原因を一つ一つ明かにして行く試験が必要である。
卵の冷蔵抵抗力と云うような問題は別の角度からの検討が必要である。
十万(238)は、日124号および支124号につき、1-2令を箱飼い、または蚕箔湿布育とする条桑、3-4令を蚕箔条桑、5令を屋外条桑で飼育した場合の掃立蟻量1g当りの卵量は普通育に比べて殆ど差がなかった(条桑区が僅かに多い)と云い、佐藤ら(825)は、これと同じ品種を、1-2令は1日1回給桑の包み育、3令を1日2回給桑の包み育、4-5令は室内条桑育とした場合、春蚕、初秋蚕ともに、蛹重、発蛾歩合、種繭1kg当り産卵量などの総べての点において、普通育に劣る成績であったと報告している。これは原蚕飼育の省力法として、稚蚕期の簡易育を試験したものであるが、条桑育そのものに不安の残っている現在ではこれを更に省力化することは無理な点があるように思われる。
先きに原蚕飼有用桑の質および量と次代蚕との関係について述べたのと同様に、最終結果だけについて論議していると、良かったと云う結果と悪かったと云う結果とは何時までも平行線を辿って、正しい比較検討の余地がない。良かった原因と悪かった原因とを明確にするための試験が現在の原蚕条桑育の問題についても最も望まれる処である。
蚕は、普通、上蔟後3日ぐらいで吐糸を終了し、脱皮して蛹になるから、上蔟後7、8日で収繭し、蚕種製造所に持込むまでには蛹期間の約3分1が経過している。蚕が上蔟して繭を作ってしまうと、農家の蚕に対する注意は、飼育中に比べておろそかになりがちである。一般に、春蚕では上蔟後の補温に欠けることが多いが、また反対に、補温したまま温度が上り過ぎても気付かないことがある。
吐糸終了から蛹の初期にかけては、生理的にも形態的にも蚕の体に重大な転換の行なわれる時期であるから、この時期の管理がなおざりにされていることは蚕種製造技術上の一つの盲点であって、不受精卵が多発した場合にその原因を調べようとしても、製造所に入ってからの保護環境はわかるが、それ以前のことは不明な場合が多い。
糸繭の目的である繭層は上蔟後3目ぐらいででき上るが、種繭の目的である卵の造成の重要な部分は吐糸終了後に始まるのであるから、農家ばかりでなく、蚕種製造者自身が十分な注意を払う必要がある。普通に種繭保護と云えば収繭後の保護であるが、実際問題としては、これだけでは不十分である。
1 上蔟・吐糸
繭層は種繭にとって最終の目的物ではないが、吐糸が十分に行なわれず体内に絹物質が残留すると、造卵数も産卵数も少なくなるから(Ⅱ1B)、吐糸・営繭の環境に注意する必要がある。多糸量系の原種には一般に残留絹が多いと云われている。過熟、若上げは避けなければならない。
繭の縦作りがよいか横作りがよいかについては、古くから論議があり、昆虫には本来縦作りの性質があるから、この本来の性質に合わせるべきであると云う説もあるが、種繭としては縦作りよりも横作りが望ましいとされている。これは、縦作りにおいては、蚕が吐糸を終えて化蛹する際に、頭部を上に向けるので、尾部が圧縮されて交尾不能の蛾や離交し易い蛾ができるばかりでなく、上蔟後の斃蚕歩合、発蛾歩合、造卵数および産卵数などにも悪影響があると云われているからである(第34表)。
| 営繭姿態 | 発蛾歩合 (%) |
蛹体尾部異常 蚕歩合 (%) |
正常蚕卵蛾 歩合(%) |
採種総蛾数による1蛾当り | 種繭1,000粒当り正常卵 | |||
| 産卵数(粒) | 不良卵歩合(%) | 造卵数(粒) | 重量(g) | 指数 | ||||
| 横 斜 縦 |
86 76 80 |
9 74 94 |
93 74 67 |
455 415 390 |
2 4 5 |
485 473 475 |
108.8 85.8 78.0 |
100 79 72 |
池田(241)は、種々な向きに繭を作らせて実験的にこれを確かめ、全部の蚕が横作りの繭を作るような廻転蔟の使用をすすめている。ただ、池田の実験は4カ年、6蚕期に亘って繰返えされてはいるが、全体を通じて未化蛹斃蚕歩合や斃蛹歩合が非常に高く、第34表の場合(昭和19年秋)には、未化蛹斃蚕歩合が横作りにおいて35.6%、縦作りにおいて48.4%にも達している。蚕の不健康なとき、縦作りと横作りとで斃蚕歩合の差の大きいときなどに卵に関する成績の差も大きく出ているようにみえるから、蚕の健康なときには産卵に対してこれほど大きな差はないものと思われる。
加藤ら(314)は、豊白、研光、支124号、銀竜および高の5品種についての比較を行なったが、造卵数は、支124号において縦作りが37.9±8.1粒少なかっただけで、他の品種においては縦作りと横作りとの間に差がなかった。発蛾歩合、正常産卵蛾歩合にも一定の傾向の違いは認められなかった(Ⅲ5)。
吐糸中の蚕は繭中で体の位置を種々に変えているが、吐糸を終えると繭の一端に頭部を向けて静止し、化蛹する。この静止するときの頭の向きは重力と光とによって強く支配され、水平に営繭させて一方向から光をあてると、光の来る方向に頭部を向けているものが、反対に向いているものより多い。しかし、光よりも重力の影響が強いので、傾斜した位置で営繭させると、光の方向とは無関係に大部分または全部のものが上方に頭部を向ける(第35表)。
| 営繭角度 | 光線 | 繭内の蛹の頭部の向き | |||
| N品種 | S品種 | ||||
| 上(頭) | 下(頭) | 上(頭) | 下(頭) | ||
| 15° | 全暗 上明、下暗 上暗、下明 |
28 28 27 |
2 2 3 |
23 27 19 |
7 3 11 |
| 30° | 全暗 上明、下暗 上暗、下明 |
30 30 30 |
0 0 0 |
29 30 25 |
1 0 5 |
| 45° | 全暗 上暗、下明 |
30 30 |
0 0 |
30 29 |
0 1 |
| 90° | 全暗 上暗、下明 |
30 30 |
0 0 |
30 29 |
0 1 |
但し、品種によって反応にかなり相違がある。また、水平に営繭させた繭の蛹の尾端に当る方の破風部に250luxの光をあてると、吐糸終了後5時間(上蔟後約63時間)では約25%の蚕が光の方向へ頭部の向きを変えたが、吐糸終了後10時間(上蔟後約68時間)では光に反応して向きを変えるものはなかった。これに対し、暗中において45°の角度で営繭させたものの上下を反対にすると(45°の傾斜で)、吐糸終了後5時間では約半分、10時間でもなお約22%の蚕が上方へ向きを変えた。即ち、250luxの光よりも45°の角度による重力の刺戟の方が強く(おそくまで)働いている(422)。これは、供試品種33×73、温度25℃ 、湿度約80%、上蔟後約20時間で営繭中の幼虫が僅かに透視できる程度に吐糸し、その後38時間ぐらいで吐糸終了、その後更に35時間で化蛹した場合における調査結果である。
池田(241)は、上蔟後3日間を第1期、次ぎの3日間を第2期、その後発蛾までを第3期として、繭の向きを変える実験を行ない、頭部を上方に向けて第2期(吐糸終了から若蛹の期間)を径過させると前記の縦作りの被害が最も著しいと云っている。縦作りによる尾部の圧縮障害は、第2期の初めに(欧16号では上蔟後102時間目)に上下を逆転すれば防止できる(小泉の結果からも理解できる)が、斃蚕を多くしたり発蛾歩合、正常産卵蛾歩合および造卵数などを低下させる生理障害は、この時期に繭を逆転しても防止できなかった。
吐糸能率を高め、残絹量を少なくするための蔟中温度は、川畑(323)によれば23℃で、25.5℃、23℃゜、20℃における残絹量はそれぞれ13%、4%および6%であった。乾繰上蔟(湿度75%)に比べて多湿上蔟(90%)、有気流上蔟(風速1.0m/sec)に比べて無気流上蔟はそれぞれ残留絹が幾分多いと云うから、換気のよい乾燥状態で上蔟させることも必要である。
2 卵母細胞および蛹の発育階程
蚕種の保護が胚の発育に基礎をおいているのと同様に、種繭の保護も卵母細胞および蛹の発育に基ずき、その生理に応じた取扱いをする必要がある。
卵母細胞の発育には、胚の場合のような一般的に採用されている基準階程がないが、小沢(720)は、実用のために七つの発育階程を区別した(第16図)。

第16図 卵母細胞の発育階程。数字は階程番号(本文参照)
1 初期球状期 栄養細胞と卵母細胞との配列が交互栄養型(Ⅰ2Aa)の特徴を現わした時期で、卵母細胞は球状であるが、栄養室側が平たく、下方に向ってまるい。その核は細胞の中央に位し、球形である。核内に大形のまるい仁がみられる。包卵細胞は未だ一定の配列を示さず、卵母細胞の外側に散在する。
2 盃状期 栄養細胞の発育が進み、卵母細胞は、これに接する側が圧迫されて、盃状を呈する。卵母細胞の核は片側に寄り、仁は染色性が淡くなる。包卵細胞は卵室外側にほぼ1列にならび、次第にこれを取囲む。栄養細胞とその上位の卵母細胞との間に橋帯ができる。
3 半球状期 包卵被膜がほぼ完成し、これに包まれた卵母細胞が濾胞のなかばを占めて半球状を呈する時期である。卵母細胞に淡黄色の脂肪性卵黄が蓄積されるために、生の材料で核を透視することができなくなる。卵母細胞の中央部に、栄養細胞からの大量の物質の流入が認められる。卵母細胞を取囲む包卵被膜細胞は柱状の丈の高い細胞になる。栄養細胞と卵母細胞との間にも、周囲から包卵細胞が入込んでくるが、なお巾の広い連絡が残っている。
4 栄養細胞退化期 卵母細胞は縦の方向に長さを増し、上部の大きく欠けた楕円形になる。栄養細胞と卵母細胞とはなお連絡し、前者から後者への物質の流入が認められる。
5 包卵被膜完成期卵 栄養細胞と卵母細胞との間に入込んだ包卵被膜細胞が発達して、卵室を完全に取囲む時期である。卵母細胞は完全な楕円形に近付き、上部に僅かに凹みを残すが、大きさはほぼ成熟卵に等しい。卵黄膜と包卵被膜との開に卵殻が形成され初める。
6 卵殻分泌期 包卵被膜細胞から卵殻の分泌される時期である。
7 完成期 卵管本部の卵が包卵被膜から離脱して下降する時期である。
蛹令別に、1卵管中に含まれる卵をこれらの階程によって示したのが第36表であるが、これに勝又(317)の蛹の発育階程を併記した。一般に、複眼が黒色になったときは、その保護条件における蛹期間のなかばを幾分過ぎた時期とされている。
| 蛹令 | 調査 卵管数 |
卵母細胞の発育階程 | 蛹の外形 | |||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 産下卵 | |||
| 化蛹 1日目 |
本 |
粒 |
粒 |
粒 |
粒 |
粒 |
粒 |
粒 |
粒 |
体柔らかく黄白色または淡黄褐色。翅下の腹部 白色。頭部透明、正中腺および触角基部黒色に 透視される。 |
| 2日目 | 11 | 81 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 翅下の腹部褐色となる。 |
| 3日目 | 11 | 39 | 59 | 8 | 0 | 0 | 0 | 0 | 0 | 体硬化、固有色 |
| 4日目 | 12 | + | 30 | 22 | 26 | 0 | 0 | 0 | 0 | 尾部透明となる。 |
| 5日目 | 10 | + | 24 | 13 | 31 | 38 | 0 | 0 | 0 | 眼着色し始め、小豆色。 |
| 6日目 | 11 | + | 16 | 24 | 51 | 0 | 0 | 0 | 眼濃小豆色。 | |
| 7日目 | 10 | + | 13 | 38 | 28 | 0 | 0 | 眼黒色。頭部白色不透明。 | ||
| 8日目 | 11 | + | 18 | 21 | 53 | 0 | 0 | 生殖キチン板、爪、肢の環節 など黒色に透視される。 |
||
| 9日目 | 10 | + | 13 | 13 | 69 | 8 | 0 | 体銹色を帯び、触角着色し始む。 | ||
| 10日目 | 12 | + | 13 | 48 | 31 | 0 | 触角暗褐色、体銹色を増し、柔かし | |||
| 11日目 | 11 | + | 12 | 17 | 67 | 0 | 体柔かくなり、土色、皺多く、翅の斑紋みえるもの あり(みえないで発蛾することもある)。 |
|||
| 発 蛾 | 20 | + | 11 | 6 | 79 | 0 | ||||
| 産卵後 | 10 | + | + | 2 | 0 | 81 | ||||
このような蛹の形態変化は複雑な変態および発育の生理に件なって進行している訳であるが、生理作用の外面的な現われの一つである呼吸は蛹期間を通じて第17図のような消長を示す。蚕の酸素消費量は5令の中期以後低下を続け、化蛹3日目頃に最低になるのであるが(266)、伊藤(280)は、蛹体休眠をする鱗翅目昆虫の生理状態と比較して、この呼吸最低期は組織崩壊と組織形成との二つの異なる代謝の転換期に当り、生理的には休眠状態に近いものと考えている。

第17図 蛹期間における酸素消費量の消長(伊藤(280))
点線は雌、実線は雄
この考えの当否は兎も角、化蛹3日目頃の呼吸最低期を中心とする時期は、蛹の保謹上からも極めて重要な転換期である。この最低期を境に雌と雄との曲線の上下が入れかわることについて、高岡(958)は、卵巣を除去すると雌の曲線が雄型になることから、雌における卵巣の発育に伴なう酸素消費の増加が原因であろうと考えている。但し、高岡の酸素消費量曲線は、化蛹後約50時間目を最低として、雄はやや直線的に尻上りの増加を示し、雌は170時間目頃に最高となり以後減少すると云うから、第17図とは少しく異なるもののようである。後に述べるように、休眠ホルモンが卵に作用し初めるのはこの最低期である。蛹期中で冷蔵に最も耐えるのはこの時期であり、これに次ぐのが、第2の呼吸低下期に当たる化蛹前日であることも注目される。
木村・水出(379)によれば、化蛹後、温度25.3℃(23.5℃-27.5℃)、湿度53.6%(41-65%)で保護した場合(蛹期間は雌平均253時間、雄平均246時間)の蛹重減耗曲線には、化蛹後80時間目前後の、複眼が淡茶色に着色し初めたときに変曲点があり、
┌ W1=4.655 e-0.0258t1 +95.25 0≦t1≦80
雌 ┤
└ W2=-0.011210(t2-70)1.3564+96.05 80≦t2≦240
┌ W1=5.619 e-0.0280t1 +96.05 0≦t1≦80
雌 ┤
└ W2=-0.004914(t2-70)1.5864+94.83 80≦t2≦240
で表わせると云う。但し、t1、t2は化蛹後の経過時間、W1、W2は、t1、t2における蛹重(化蛹直後を100とする指数)である。品種はC110×N112であった。
3 蛹の保護
A 蛹の発育
a 保護環境と蛹期間
普通の保護温度の範囲内では多くの場合、雌よりも雄の蛹期間が短い。蛹期間は温度の上昇に連れて短かくなるが(第37表)、堀内ら(223)によれば、温度に正しく比例する訳ではなく、20℃と25℃との間では40%以上も短縮するが、25℃と30℃との間の短縮は10%余りに過ぎない。温度による蛹期間の変化は、また雌雄によって異なり、雌雄差は25℃において最大で、30℃、20℃および15℃においてはこれよりも差が小さい。特に、15℃においては、従来から知られているように雄よりも雌の蛹期間が短かくなる傾向がみられたが、30℃においても、幾分これと同じような傾向のあることがわかった(第40表)。 この関係は品種によっても異なる。
| 保護温度 ℃ |
性別 | 蛹 期 間 (日) | |||
| 日122号 | 日124号 | 支115号 | 支124号 | ||
| 30 | ♀ ♂ |
**10.6 10.0 |
** 9.6 9.3 |
* 8.0 7.7 |
** 8.3 7.8 |
| 25 | ♀ ♂ |
**11.9 11.1 |
**10.6 10.3 |
9.4 9.1 |
* 9.8 9.5 |
| 20 | ♀ ♂ |
**20.3 19.8 |
18.5 18.6 |
15.9 **16.9 |
16.7 16.7 |
| 15 | ♀ ♂ |
26.3 26.0 |
24.6 24.9 |
22.1 22.3 |
**23.0 22.4 |
上蔟は殆ど総べての品種において雄が雌より早いが、化蛾は必ずしもそうではなく、諸星・小林(599)によれば、個体別に化蛹時刻を調べ、24℃と19℃とで保護したものについて蛹期間の平均を求めると、両温度区を通じて、カムボージュ、欧16号、欧19号などにおいては雄が雌より短かく、大造、日112号などでは雌が雄より短かかった。温度との関係については、保護温度の高いほど雄の相対的発育速度が雌より大きくなり、温度が低いほど雌の相対的発育速度の大きくなる傾向がある。これは伴性晩性遺伝子の働きとホルモン作用の拮抗とによって説明できると云う(597)。永友(645)、室賀(617)も蛹期間の雌雄差についての遺伝学的説明を試みている。
堀内ら(223)の上記の実験は、6時から21時までを明保護、21時から翌朝6時までを暗保護にして行なったのである.が、6時に明るくすると大部分の蛾は3時間以内に発蛾した。その後にも不時発蛾するものがあるが、不時発蛾は、試験した温度の範囲内(第37表)では、25℃保護において最も少なかった。
保護温度の高低によって蛹期間が伸縮すると、蛹の外形の発育階程もこれに連れてほぼ均等に伸縮するもののようで、堀内らの試験の範囲内では、複眼小豆色のときが総ての保護温度を通じて蛹期間の中点であった。
蛹体の外観によって発蛾までの日数を推定することは古くから行なわれており、一般に、複眼が黒色になったときは蛹期のなかばを過ぎた時期であると云われているが、最近の品種についての加藤(311)の調査を第38表に示す。
| 蚕品種 | 日122号 | 秋花 | 支115号 | 支124号 | ||||||||
| 蛹の外観 | 22℃ | 25℃ | 28℃ | 22℃ | 25℃ | 28℃ | 22℃ | 25℃ | 28℃ | 22℃ | 25℃ | 28℃ |
| 上蔟 吐糸終了 化蛹 複眼着色始む 複眼淡黒色 複眼濃黒色 生殖外器着色始む 生殖外器黒色 生殖キチン板透視 触角濃黒色 蛹皮土色 |
19日 15 13 10 9 7 6 5 4 2 |
18日 15 13 10 9 7 6 5 4 2 |
15日 12 11 8 6 5 4 3 2 |
20日 15 14 11 7 6 5 3 2 |
17日 13 12 10 6 5 4 3 2 1 |
15日 12 11 9 5 4 3 2 1 |
16日 13 12 10 7 5 3 2 1 |
14日 11 10 8 6 4 3 2 1 |
13日 11 10 7 5 3 2 1 |
16日 13 12 9 6 4 3 2 1 |
16日 13 12 9 6 4 3 2 1 |
14日 11 10 8 6 4 3 2 1 |
保護湿度60-90%の範囲内では、湿度の低いほど蛹期間は短かく(第39表)、裸蛹は繭中のものより蛹期が短かいと云う。
| 湿 度 | 平均蟄繭日数* |
| 60% 70 80 90 |
13.2日 13.7 13.7 14.1 |
b 化蛹までの蔟中保護温度と蛹期間
化蛹後の保護温度と蛹期間との間に上記のような関係があるのに対し、化蛹までの蔟中保護温度を25℃にしたものと30℃にしたものとの蛹期間を比較すると、後者が前者よりも常に長いと云う(第40表)。
牛込(1127)は、日1号、日110号、支4号、支102号および欧7号について、上蔟以後の保護温度78-83゚F(25.6℃-28.3℃)の範囲内においては、温度の高いほど蛹期間が短縮するが、85゚F(29.2℃)また87゚F(30.6℃)がその限界で、保護濃度がそれ以上になると蛹期間は却って延長すると述べている。しかし、これは化蛹前および化蛹後を含めた期間であって、両者を区別した期間には言及していない。
| 化蛹後の保護温度 (℃) |
化蛹までの蔟中温度 (℃) |
性別 | 蛹期間(日) | |
| 日124号 | 支124号 | |||
| 30 | 25 | ♀ ♂ |
10.9 10.7 |
9.0 8.8 |
| 30 | ♀ ♂ |
11.0 *11.4 |
**9.4 8.7 |
|
| 25 | 25 | ♀ ♂ |
**13.7 12.6 |
**11.2 10.6 |
| 30 | ♀ ♂ |
13.9 13.4 |
**11.5 10.5 |
|
| 20 | 25 | ♀ ♂ |
20.8 20.9 |
**18.0 17.2 |
| 30 | ♀ ♂ |
21.3 21.2 |
**18.8 17.3 |
|
| 15 | 25 | ♀ ♂ |
30.5 30.5 |
**26.8 26.1 |
| 30 | ♀ ♂ |
31.9 31.0 |
**27.8 26.6 |
|
c 発育段階別保護温度と蛹の発育
伊与田・米山(295)は、蛹期の卵巣重にみられる3段階の発育(Ⅰ2Abi)は蛹の外形変化と関連があり、発育の前期は化蛹後、複眼の着色し初める時期まで、中期はその後触角の着色し初める時期までであるとし、低温(15℃)に保護すると前期と後期の延長は著しいが、中期の延長は比較的少ないと述べている(第41表)。
| 蛹令 (日目) |
第1区 全期高温 | 第2区 前期高温、中期 低温、後期高温 |
第3区 前期低温、 中・後期高温 |
第4区全期低温 | ||||||
| 蛹重 (g) |
卵巣重 歩合(%) |
蛹重 (g) |
卵巣重 歩合(%) |
保護 温度(℃) |
蛹重 (g) |
卵巣重 歩合(%) |
保護 温度(℃) |
蛹重 (g) |
卵巣重 歩合(%) |
|
| 化蛹当日 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 |
8.6 - 8.12) 8.23) 8.06) 7.78) 8.09) 8.110) 5.511) |
0.7 - 4.5 8.5 20.0 33.0 38.1 37.3 47.5 |
8.6 - 8.1 8.2 8.05) -6) - 8.17) - - - 8.1(5)9) - 4.711) |
0.7 - 4.5 8.5 13.2 - - 26.4 - - - 33.1 - 49.6 |
30 〃 〃 〃 15 〃 〃 〃 17 19 21 30 〃 〃 |
8.6 - - 8.6 - - 8.5 - - 8.3 7.96) 7.4(5) 7.6 4.911) |
0.7 - - 2.0 - - 3.3 - - 6.5 21.0 37.0 33.6 49.5 |
30 〃 〃 〃 15 〃 〃 〃 17 19 30 〃 〃 〃 |
8.6 - - 8.6 -1) - 8.5 - - 8.33) -4) - - 7.5 - - 7.99) - - 8.3(5) - 5.8(10)11) |
0.7 - - 2.0 - - 3.3 - - 6.5 - - - 31.7 - - 33.7 - - 31.2 - 46.1 |
加藤(311)は、全期間22℃、25℃および28℃の恒温区と、複眼黒色期(第38表の淡黒色期と濃黒色期とを合わせた時期)以後を変温した区とを設けて、上蔟後、発蛾までの保護を行ない、発蛾および産卵におよぼす温度の影響を比較した(第42表)。これによると、発蛾歩合は、上蔟後、発蛾までの全期間を同一温度で通した場合には28℃区が最も悪く、牛込(1127)の成績において、上蔟後を68゚F(20℃)、73゚F(22.8℃)、78゚F(25.6℃)および83゚F(28.3℃)に保護した場合に83゚F区の最も劣っていたことと一致した。22℃区と25℃区との成績は、品種的な関係があるためか、優劣をきめかねる結果であるが、複眼黒色期以前を22℃、以後を25℃に保護した区(表の22℃→25℃)が全期間25℃の区に勝っているのをみると、少なくとも発蛾歩合に関しては、化蛹前あるいは蛹初期の保護の適温は25℃以下のように考えられる。
| 蚕品種 | 保護温度 (℃) |
発蛾 歩合(%) |
正常産卵蛾 歩合(%) |
正常産卵蛾についての調査 | |||||
| 1蛾当り総 産卵数(粒) |
正常卵 歩合(%) |
卵長 (mm) |
卵巾 (mm) |
1g卵 数(粒) |
100粒 卵重(g) |
||||
| 日122号 | 22 25 28 22→25 22→28 25→22 28→22 |
88 83 67 86 78 83 70 |
86 86 57 76 76 88 47 |
558 542 462 612 525 517 526 |
94 96 87 96 96 95 92 |
1.29 1.29 1.33 1.30 1.29 1.31 1.34 |
1.04 1.05 1.07 1.05 1.04 1.06 1.09 |
1,731 1,728 1,591 1,698 1,683 1,685 1,637 |
0.0578 0.0579 0.0629 0.0589 0.0594 0.0594 0.0611 |
| 秋 花 | 22 25 28 22→25 22→28 25→22 28→22 |
77 63 55 70 68 68 66 |
70 69 47 55 40 51 40 |
511 494 457 497 514 468 445 |
97 95 78 96 96 92 73 |
1.36 1.34 1.37 1.33 1.33 1.34 1.36 |
1.10 1.09 1.10 1.08 1.08 1.08 1.11 |
1,676 1,667 1,615 1,671 1,646 1,680 1,617 |
0.0597 0.0600 0.0619 0.0598 0.0608 0.0595 0.0618 |
| 支115号 | 22 25 28 22→25 22→28 25→22 28→22 |
84 92 61 94 69 96 69 |
58 88 53 81 60 72 63 |
503 559 521 550 558 505 530 |
94 94 93 92 93 93 93 |
1.31 1.33 1.38 1.35 1.37 1.37 1.40 |
1.05 1.05 1.09 1.07 1.08 1.07 1.10 |
1,821 1,832 1,680 1,794 1,761 1,724 1,673 |
0.0549 0.0546 0.0595 0.0557 0.0568 0.0580 0.0598 |
| 支124号 | 22 25 28 22→25 22→28 25→22 28→22 |
90 91 74 92 73 92 62 |
66 88 53 68 65 73 60 |
520 563 536 612 503 565 469 |
93 94 88 92 92 94 90 |
1.27 1.28 1.30 1.27 1.26 1.29 1.29 |
1.07 1.07 1.08 1.06 1.05 1.08 1.09 |
1,770 1,768 1,700 1,787 1,706 1,714 1,657 |
0.0565 0.0566 0.0588 0.0560 0.0586 0.0583 0.0604 |
B 保護温度と造卵および産卵
a 造卵数・産卵数
普通の種繭保護温度の範囲内では、一般に、温度の低い場合に造卵数が多いが、第43表において、日110号および昭和A(生種)は20℃保護の造卵数が25℃あるいは30℃保護に比べて多いのに対し、昭和A(黒種)は20℃保護が25℃および30℃保護よりも却って造卵数が少ないのをみると、造卵数を増加させる低温の限度は品種または条件によって相違するものと考えられる。
| 保護温度 (℃) |
日110号 | 昭和A(黒種) | 昭和A(生種) | |||
| 調査蛾数 (蛾) |
1蛾平均 造卵数(粒) |
調査蛾数 (蛾) |
1蛾平均 造卵数(粒) |
調査蛾数 (蛾) |
1蛾平均 造卵数(粒) |
|
| 30 25 20 15 |
102 270 |
519 572 |
85 40 60 33 |
615 643 602 452 |
367 505 425 |
431 497 499 |
蛹の保護温度が造卵数の増加に影響する時期は、伊与田らによって、主として先に述べた前期と考えられている(第44表)。
| 蛹の保護条件 | 調査蛾数(蛾) | 平均造卵数(粒) |
| 全期間高温(第41表第1区) 前期高温、中期低温、後期高温(同表第2区) 前期低温、中・後期高温(同表第3区) 全期間低温(同表第4区) |
7 7 7 10 |
538 528 553 599 |
しかし、この造卵数の増加は卵巣の発育とは別で、卵巣の重量増加に最も大きく影響するのは中期の高温であると云う。
永井(630)が、支17号を65゚F(18.3℃)、75゚F(23.9℃)および90゚F(32.2℃)に分けて上蔟させ、化蛹後は何れも70゚F-72゚F(21.3℃-22.2℃)で保護した結果によると、卵管1本当りの完成卵数が65゚F区83.6粒、75゚F区94.4粒、90゚F区79.7粒で、75゚F区が最も多かった。これからみると、化蛹前の保護温度も造卵数に影響している。
造卵数は、このように、ある程度までは、蛹期の保護温度の低いほど増加するが、造成された卵の産卵歩合、正常産卵蛾歩合、不受精卵歩合などからみると、後に述べるように、低温保護の成績が不良で、卵の孵化にも悪影響があるから、造卵数だけによって保護温度を決定することはできない。
地方蚕業試験場が協同で行なった収繭後の保護温度試験においては(1147)、支106号、日110号、日7号、支14号、欧16号、欧18号、支105号その他の品種につき、25℃、23℃、20℃および15℃保護を比較したが、産卵数の最も多かったのは23℃-25℃保護で、これに次ぐのが20℃保護であった。30℃および15℃に保護したものは産卵が著しく少なかった。
23℃-25℃においては不受精卵も最も少なかった。永井(626)の試験においても、20℃、24℃および27℃のうちでは24℃が最もよかった。
農林省蚕糸試験揚が最近の蚕品種について試験した成績をみても(802)、発蛾歩合、正常蚕卵蛾歩合、産卵数などを綜合して、種繭保護の適温は23℃-25℃の範囲にあり、やや委しく云えば
日122号 24℃-25℃ 支115号 25℃
日124号 23℃-24℃ 支122号 23℃-25℃
日125号 〃 支122号(太) 〃
日502号 〃 支124号 〃
支25号 23℃ 支503号 〃
であった、と結論されている。第42表においても、正常卵蛾歩合は蛹を25℃またはその付近の温度に保護した場合に最も高い。。足立ら(13)は産卵量および産卵速度からみて、23℃が最もよかったと報告している。
平山・館石(202)は上蔟後を30℃、24℃、および18℃に保護する3恒温区と、1)化蛹まで、2)化蛹から腹眼着色まで、3)複眼着色から触角着色まで、4)触角着色から発蛾まで、の期間それぞれ毎日、日中(8時から20時まで)30℃、夜間(20時から8時まで)18℃、平均24℃で保護する4変温区とに分けて比較し、第45表に示す結果を得た。
| 試験区 | 春蚕期 | 初秋蚕期 | ||||||
| 調査蛾 数(蛾) |
産卵数 (粒) |
受精卵 歩合(%) |
調査蛾 数(蛾) |
造卵数 (粒) |
産卵数 (粒) |
受精卵 歩合(%) |
産卵歩合 (%) |
|
| 30℃区 24℃区 18℃区 変温1区 変温2区 変温3区 変温4区 |
16 16 14 14 14 10 14 |
494 586 456 571 552 473 585 |
90 98 47 99 95 91 98 |
21 23 21 14 21 21 15 |
484 565 464 550 511 499 570 |
400 556 364 516 483 460 533 |
94 98 62 98 98 97 97 |
82 98 78 93 94 92 93 |
これによると、18℃恒温区が最も悪い。また、平均温度は同じ24℃でも、恒温区に比べて変温区が劣り、変温区相互の間では第3区が悪かった。これは伊与田らが卵巣の発育に対する蛹期保護温度の影響の最も大きい時期としている中期の変温に相当する。造卵数は、伊与田らの結果とは反対に、18℃恒温区が最も少なく、24℃恒温区が最も多く、、第43表の昭和A(黒種)と同様な結果を示した。卵の大小については言及されていないが、1g蟻数が
30℃区 2285頭 変温1区 2562頭
24℃区 2556 〃 2区 2678
18℃区 2895 〃 3区 2683
〃 4区 2632
であったのをみると、卵は、後に述べるように、保護温度の高いほど大きかったものと考えられる。変温区は総べて、24℃恒温区に比べて、1g蟻数が多い(卵が小さい)から、この点からみると、平均温度としては同じでも、変温区は恒温区に比べ、生理的には低目な温度として作用したように思おれる。
1g蟻数の多かったものほど、一般に、蟻蚕の絶食生命時数は短かった。
正常産卵蛾の産卵数は、発蛾したもののうち、産卵数について選抜されたものの産卵数であるから、各区間の差の小さくなるのは当然であるが、第42表において、発蛾歩合や正常産卵蛾歩合に比べ、各区間の差が小さい。特に、後半28℃保護のもの(22℃→28℃)が25℃恒温のものに非常に接近した成績を示しているのが注目される。牛込(1127)が、産卵数は、上蔟後、発蛾までの保護温度68゚F(20℃)または73゚F(22.8℃)のものが、78゚F(25.6℃)または83゚F(28.3℃)のものよりも多いが生殖外器キチン板を透視し得るようになった時期(第36表)以後は、83゚Fに保護しても産卵数が少なくならなかつたと云っているのもこれと同じ傾向を示すものと考えられ、蛹の後期には産卵敬に対する高温の悪影響が減少するものと考えられる。
永井(631)によれば、産絹量の多い系統の蛹は、保護温度21-27℃(湿度75%)の範囲内においては温度の高いほど、湿度63-99%(温度24.5℃)の範囲内では湿度の低いほど産卵歩合(造卵数に対する)が低下したが、これは産絹量の多いものほど吐糸による体水分の減少の多いことに関係があるものと考えられると云う。但し、蛹は裸蛹で保護した。
b 保護温度と卵の大小
蚕の卵は品種によって大きさが異なり、概して、欧州種は支那種および日本種に比べて大きく、日本種の卵は欧州種および支那種に比べて長目である。卵の大きさは、また、蚕の栄養状態によって相違する(Ⅱ2Ac)ほか、同じ蛾の卵でも、初めに産むものは大きく終りのものは小さいが(242,1142)、更に、種繭の保護温度によって相違し、温度の高いほど卵は大きく、1g卵数は少なく、100粒卵重が大きい(第42表)。
木暮・山本(408)によれば、保護温度の高まるに連れて、卵が大きくなると共にその長巾率が小さくなり、卵はまるみを増してくる。宮坂(568)も同様な傾向を報告している。種繭の保護温度が卵の大小に影響する時期は主として化蛹から複眼黒色期までの間で、その後の影響は皆無ではないが少ないと云うから、保護温度が造卵数に影響する時期とほぼ一致している。牛込(1127)は、化蛹前の温度もこれに影響するらしい成績を得ている。
高温保護によって卵の大きくなる理由について、木暮らは、1)包卵被膜細胞の生理作用が増大するのか、2)高温のために発育を中止する卵が生じ、残った卵がその養分を取り込かのか、二つのことが考えられるが、温度の高くなるに連れて卵形の大きくなるのに対して、卵数の変化は温度の上昇と無関係であるから、卵形の大きくなることと造卵数の多少とは無関係であって、上記第2の説明は実際に合わないと結論している。
卵数の変化が温度の上昇と無関係であると云うのは、第43表の昭和A(黒種)の場合を指しているのであるが、これは、卵数を増加させる低温の範囲には、それぞれの場合に応じて限度のあることを示すものとも考えられるから、これだけでは卵数の多少と卵の大小との関係を否定する根拠として不十分なように思われる。卵数が減少してその養分が残りの卵に利用されるのは、氏らの考えているように退化卵からの直接の養分吸収ではなく、発育すべき卵数が制限せられ、配分する養分に余裕が生じ、この分が間接に利用されるものであろう(Ⅰ2Abi)。また卵形に対する温度の影響の最も大きい化蛹から複眼黒色期までの時期は卵巣発育の前期から中期の初めに当り、卵巣は未だ絶対重が小さく、発育も緩徐なときであって、顕著な卵巣の発育は複眼黒色期以後である(第41表)。これらのことと、保護温度が卵の大小に影響する時期と造卵数に影響する時期との一致していることを考え合わせると、卵の大小におよぼす種繭保護温度の影響は2段階に分けて検討すべきもののように思われる。即ち、第1段階として、蛹の初期に温度の直接あるいは直接に近い影響によって、発育すべき卵の数、従って養分配分のプランがきまり、第2段階として、複眼着色期以後の卵の発育によってこのプランが実現されるのであって、卵数の多少と卵の大小とは間接的に関係があるものと考えるべきであろう。
種繭の高温保護によって大きく作られた卵は、催青日数が長く、蟻蚕体重も重いから(第46表)、卵殻ばかりではなく、卵黄の増加していることが明かである。
| 蚕品種 | 保護温度 (℃) |
蟻蚕重量 | 催青日数 | ||
| 調査頭数(頭) | 1頭平均(mg) | 調査蛾数(蛾) | 平均孵化日数(日) | ||
| 昭和A (生種) |
30 25 20 15 |
2,352 2,351 1,017 894 |
0.4252 0.3964 0.3752 0.3365 |
30 - 30 - |
10.2 - 9.5 - |
| 昭和A (黒種) (人工孵化) |
30 20 |
1,725 1,941 |
0.4637 0.4174 |
20 20 |
10.9 10.2 |
直接の実用性は兎も角、造卵数増加の問題の主要項目の一つである卵の数と量との関係の転換(Ⅰ2Abi)が、種繭の保護温度の管理によって一部達成されることは興味がある。
上蔟後の保護温度についてこれまで述べてきた処を綜合すると、全期間恒温の場合には23-24℃、前期と後期との保護温度を区別する場合には、複眼黒色期頃を境にして、前期をやや低目、後期をやや高めにするのがよいと考えられる。産卵が少なく、異常卵の多い傾向のある日122号の蛹を、前期やや低温、後期やや高温に保護して好成績をおさめた例を第47表に示す。但し、不受精卵や催青死卵については、種繭の保護温度以外にも原因の多いことを考慮する必要がある。
| 飼育場所 | 保護温度 | 造卵数 (粒) |
産卵数 (粒) |
異常卵 | ||
| 不受精卵 (粒) |
催青死卵 (粒) |
合計 (粒) |
||||
| A | 1.全期22℃ 2.前期22℃、後期26℃ 3.前期26℃、後期22℃ 4.全期26℃ |
506 520 463 478 |
453 460 424 438 |
22 18 20 23 |
10 8 12 15 |
42 41 38 43 |
| B | 1.全期22℃ 2.前期22℃、後期26℃ 3.前期26℃、後期22℃ 4.全期26℃ |
654 650 590 582 |
589 597 542 540 |
21 22 25 29 |
12 12 15 16 |
51 50 54 50 |
| C | 1.全期22℃ 2.前期22℃、後期26℃ 3.前期26℃、後期22℃ 4.全期26℃ |
510 502 432 440 |
443 439 418 425 |
18 15 20 25 |
9 10 13 12 |
45 41 39 44 |
c 保護温度と卵の越年性
種繭の保護温度と卵の越年性との関係については、渡辺(1139,1141)によって、高温保護は不越年性卵を多くすることが報告されていたが、最近、休眠ホルモンを注射した不越年卵性蛹(蛾になってから不越年性卵を産む蛹の意味である。以下これに準ずる。)は、注射後を高温におくよりも、15℃
に5日間ぐらい保護した方が越年性卵を多く産むことがわかって(165,169)、ホルモン作用の面からも証明された。
しかし、この渡辺の報告は、二化性または四化性種の卵を20℃前後の中間温度で催青した場合の蚕を飼育して得た蛹の保護温度について述べたもので、普通のように二化性種を高温(25℃)で催青した場合には、上蔟後の保護温度がかなり高いときにも不越年性卵は出にくい(第48表)。
| 催青温度 | 4令期以後の保護温度 | 産卵結果 |
| 25℃以上 | 飼育中25℃以上、上蔟後30℃ 〃 17℃ 、 〃 20℃ |
越年卵 〃 |
| 15℃以下 | 飼育中25℃以上、上蔟後30℃ 〃 17℃ 、 〃 20℃ |
不越年卵 〃 |
| 20℃前後 | 飼育中25℃以上、上蔟後30℃ 〃 17℃ 、 〃 20℃ |
越年卵と不越年卵(不越年多し) 〃 (越年多し) |
この表には、4令期以後の飼育温度17℃の場合も示してあるが、これでわかるように、二化性種を25℃で催青した場合には、普通には、飼育温度は稚蚕期、壮蚕期とも化性に影響することが少ない。
渡辺(1141)は、産卵の越年現象を、卵の発育を一時的に抑制するある物質の存在によって起こるものと仮定し、この物質を抑制質(inhibitory
substance)と名什けた。この説明によると、二化性種を高温で催青した場合には、胚の体中に多量の抑制質が生成し、これが蚕児、蛹を径て次代の卵に伝わり、その卵を越年させるが、低温で催青すると抑制質が生成されないため、次代の卵が不越年性になるのである。このように催青温度が高温または低温であると、卵の越年性または不越年性は安定しており、上蔟後の温度によって影響を受けないが(第48表)、中間温度で催青すると、抑制質の生成される胚と生成されない胚とが生じ、生成されたものにおいても、抑制質の量は個体間に変異がある。抑制質の少い蚕は、4令期以後、卵巣内で卵の分化の始まる頃からの保護温度の影響により、温度が比較的低いときには、少量の抑制質によってもその卵巣内の卵を越年化させることができるが、温度が高いと抑制質の作用が発現し得ず、卵は不越年性になると云うのである。
上蔟後の保護温度が低いときに再出卵の多いことも、同様に抑制質の仮定によって説明されている(1143,1144)。ここに再出卵と云うのは、漿膜が着色して越年性卵の外観を呈する卵でありながら、産卵後2週間前後で孵化し、1蛾の卵の中に越年性卵と混在しているもののことである。
中間温度で催青した抑制質の少ない蚕を、上蔟後、中間温度で保護すると、抑制質の作用がある程度働くために、完全な不越年性にはならずに産み出されるが、越年性が不安定な状態にあるため、産卵後の保護温度が高いと再出卵になる。これに対して、上蔟後の保護温度が高温であると、抑制質の少ない蛹の卵は不越性卵になることが多く、不安定な状態で産み出されるものが少ない。従って、再出卵も少ない。即ち抑制質の少ない卵が、保護温度の相違によってあるいは再出卵になり、あるいは不越年性卵になるのであるから、不越年卵蛾区と再出卵蛾区との合計は、上蔟後の保護温度が高くても中間温度でも、殆ど変わりがないことになる(第49表)。高温催青を行なえば、この場合の再出卵は殆どあるいは全く生じない。
| 試験 | 上蔟後発蛾 までの温度(℃) |
総蛾区数 (蛾) |
生種蛾区 数(蛾) |
再出卵蛾 区数(蛾) |
(生種蛾区+再出卵蛾区)/総蛾区数×100 (%) |
| A | 30 20 |
144 227 |
53 15 |
20 115 |
50 57 |
| B | 30 20 |
122 216 |
67 71 |
7 49 |
60 55 |
抑制質による越年性の決定は、産卵後、低温に保護すると数日中に行なわれるから、産卵後12時間以内に15℃に移して3日間保護すれば、その後は高温に戻しても再出卵は殆ど発現せず、15℃に5日間おけば全く発現しないのが普通であると云う(Ⅴ2B)。
これに対し、木暮(406,409)は、蛹の保護温度が高いと着色卵中の不越年卵(再出卵)歩合は減少し、着色卵が再出卵になるか否かは催青中の温度および光線とは無関係であると考えた(Ⅴ2A)。室賀(610,616)は、木暮と同様に、催青温度と着色卵中の再出卵歩合との間には関係のないことを認めたが、上蔟後、発蛾までの温度が高いと、着色卵蛾歩合が減少して不着色卵蛾歩合の増加すること、および不着色卵中の越年性卵歩合が高まると云う結果を挙げて、蛹の高温保護が再出卵を少なくするようにみえるのは、抑制質の少ない越年性の不安定な着色卵蛾が不着色卵蛾に変わり、残った着色卵蛾の産む卵には再出卵が少ないためであって、本質的に越年性卵を増加させるのではないと云い、越年性と着色性とは共に催青中に決定されて、互に密接な関係があるものと考えている。
河合(355)は、大造系統の材料を用い、25℃で催青し、蛹期を20℃と30℃とに分けて保護したが、後者に再出卵が多く、3・ヒドロキシキヌレニンの卵内移行量からみても越年性の不完全な卵であった。これは、渡辺の場合と異なり、25℃催青であるから、休眠ホルモンの分泌は十分であり、蛹期間を20℃で保護すればその越年性が一層完全になるのに対し、30℃保護においては、高温のために越年性物質(抑制質)および着色物質の卵内への移行が少なく、そのために不完全な越年性卵が生じ、再出卵になったものと解釈している。
一般に再出卵と呼ぶものの中には、上記のものとは性質の異なったものが合まれているが、これについては後に述べる(Ⅶ1A)。
ここで注意しなければならないのは、15℃に保護すると越年性が決定するとは云っても、長く15℃におき、高温(例えば25℃)に戻さなければ、越年性はいつまでも完成しないことである。
155℃に保護し続けると、60日たっても、その越年性(活性化の難易および呼吸の青酸阻害からみた)は25℃保護の産卵後2日目ぐらいのままに止まり、越年性卵ではあるが非常に活性化し易く(785)、安全な蚕種保護を行なうことができない。大野(767)によれば、15℃に15日間おいて25℃に戻したものは、翌年の孵化歩合が著しく悪かった。 これは越年性が不完全で、早く活性化する卵ができるためである。
10日までは異常がなかったと云うが、必要な日数の後には早く25℃に移して休眠の完成をはかる必要がある。
牛込(1127)は、保護温度を上蔟後の時期別に変化させて行なった試験の結果から、化蛹までを83゚F(28.3℃)に保護すると、この期間を68゚F(20℃)に保護した場合に比べて不越年性卵の多い傾向があると云っているが(第50表)、これは休眠ホルモンの作用に温度の影響する時期とは少しずれている(Ⅴ2C)。
| 蚕品種 | 保護温度(゚F) | 産卵蛾数 (蛾) |
不越年卵混産 蛾歩合(%) |
不越年卵蛾 歩合(%) |
不越年卵 歩合(%) |
||
| 前期 | 中期 | 後期 | |||||
| 日110号 | 68 73 78 83 68 68 83 68 83 83 |
68 73 78 83 68 83 68 83 68 83 |
68 73 78 83 83 68 68 83 83 68 |
173 176 244 155 179 208 198 199 211 222 |
0 0.6 0 0.7 0 0 1.1 0 0 1.0 |
0 0 0.4 3.3 0.6 1.5 1.1 1.6 3.8 0.5 |
0 0.1 1.0 4.0 0 1.2 1.0 0.6 4.0 0.6 |
| 支102号 | 68 73 78 83 68 68 83 68 83 83 |
68 73 78 83 68 83 68 83 68 83 |
68 73 78 83 83 68 68 83 83 68 |
126 147 151 186 105 128 112 105 110 162 |
0 3.7 0 1.4 1.0 0.9 9.4 1.0 0 0.6 |
2.7 4.4 2.7 8.5 2.0 2.7 0.9 0 6.4 7.5 |
1.7 4.7 2.8 8.4 1.9 2.5 5.5 0.5 6.6 5.1 |
d 保護温度と孵化の良否
i 越年種 高梨・松尾(957)が支109号を用い、上蔟から発蛾までの間を、28℃、25℃および20℃の3区に分けて保護し、採種後は何れも25℃に保護しておき3、10、20、30、50、70、90、120、150、180および210日後に5℃に移し、所定の日数毎に出庫して25℃で催青した結果の一部を第51表に示す。
これによると、90%以上の孵化歩合の続いた冷蔵期間は、上蔟後を25℃に保護したものが最も長く、20℃保護がこれに次ぎ、28℃保護が最も短かかった。また各冷蔵日数毎の孵化歩合も一般に25℃区が最もよく、これに続いて20℃区、28℃区の順位であった。表に示したのは総孵化歩合であるが、実用孵化歩合の順位もこれと同じであった。高梨らは、このような結果の得られた理由には触れていないが、蚕種保護上の重要な問題を含んでいる。
| 冷蔵日数 (日) |
高温日数 | |||||||||||||||||
| 3日 | 10日 | 30日 | 90日 | 180日 | 210日 | |||||||||||||
| 上蔟後の温度 | ||||||||||||||||||
| 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | 25℃ | 20℃ | 28℃ | |
| 30 50 60 70 80 90 100 120 140 160 180 200 220 240 260 280 300 325 |
4% 75 96 97 97 97 96 98 97 96 98 93 91 78 56 30 7 - |
8% 86 92 92 95 91 93 96 97 97 97 90 77 39 13 2 6 0 |
% 61 86 86 91 89 88 90 90 90 91 83 83 65 26 8 2 - |
0% 20 70 94 96 96 93 94 97 97 96 96 92 88 73 44 22 7 |
0% 23 89 91 93 92 93 95 97 97 93 94 91 81 54 32 9 1 |
0% 27 44 76 89 94 91 91 93 97 93 86 89 67 44 17 3 1 |
0% 21 62 94 95 96 96 97 97 96 97 96 95 90 82 69 48 11 |
% 25 62 91 95 95 96 94 95 98 97 96 93 85 68 34 16 5 |
0% 14 43 75 93 93 92 93 92 92 91 93 88 88 75 56 34 19 |
0% 4 16 44 73 84 91 95 95 96 81 89 83 73 46 37 18 7 |
0% 4 14 51 73 86 90 95 92 92 92 89 71 55 38 23 14 3 |
0% 1 17 51 68 83 91 93 94 93 90 82 85 67 59 43 23 8 |
0% - 54 72 80 85 88 92 89 80 69 59 - - - - - - |
0% - 39 57 65 74 78 81 77 70 57 43 - - - - - - |
0% - 48 63 84 80 86 88 88 78 55 59 - - - - - - |
0% - 78 86 86 82 84 76 72 61 46 20 - - - - - - |
0% - 61 76 63 51 53 54 39 35 21 12 - - - - - - |
0% - 75 77 78 75 74 69 62 45 34 19 - - - - - - |
まず、90%以上の孵化歩合を示し初めたときの孵化歩合および冷蔵日数から考えると、25℃区の活性化が20℃区よりも早いと判断されるのであるが、高温期間50日頃までは、20℃区は25℃区に比べて、活性化初期(孵化の始まる最短冷蔵日数)の孵化歩合が僅かながら殆ど常に高く、冷蔵期間が長くなって孵化歩合の低下し初める状態をみると20℃区が25℃区よりも常に早い。これは、20℃区には活性化の特別に早い卵が少数含まれていることを示すような成績である。
28℃区は活性化初期の孵化歩合が3区の中で最も低く、また孵化歩合が90%に達するまでの冷蔵日数が最も長い。これは活性化のおくれていることを示す成績のように思われる。
20℃区と28℃区とを比較すると、高温期間の短い間は、極端に冷蔵期間の長い場合を除けば、常に20℃区の孵化歩合が高いが、高温期間150日頃を境に逆転して20℃区の孵化歩合が低くなる。この時期においても、孵化歩合の最も高いのは25℃区であるから、20℃区は、25℃区および28℃区に比べて、高温期間の持続によって孵化機能を失い易い卵を合んでいるもののようである。これが、20℃区に抵抗力の弱い卵が合まれているためか否かは、与えられた数字だけからは不明であるが、20℃区には、25℃区および28℃区に比べて生理的に不安定な状態の卵の多いことは確かであろう。
20℃区に活性化の早い卵のできることは、あるいは、前項で述べたように、抑制質の少ない卵が不安定な越年性のまま残っているためであると考えら)れないこともないが、28℃区に活性化のおそい卵のできることは、休眠の成立についての渡辺の説明だけでは理解の困難な現象のように思われる。
滝沢・勝野(1005)は、上蔟後の保護温度25℃、20℃および15℃について、上と同様な方法で比較を行ない、25℃区の孵化が最もよく、15℃区の孵化の最も劣ることを観察し、また、蛹期間を前、中、後の3期に分け、25℃と15℃とを組合わせて保護した結果では、前、中期を25℃で保護したものの孵化が最もよく、この時期を15℃で保護したものは最も悪かったと述べているが、前、中、後期の分け方は不明である。
以上は人工越冬によって卵を活性化させて比較した孵化試験であるが、普通に越年種の保護をした場合にも、上蔟以後の温度によって翌春の孵化状態に相違ができる。一例として、昭和37年度関東地区蚕業試験場協力試験の結果を挙げる(印刷発表されていない)。
試験時期は春蚕期で、分場農家が上蔟後何時頃まで補温を行なうかを想定して、次ぎの3区を設けた。
A 上蔟後まる3日間を25℃目標で保護し、以後収繭まで自然温度で保護
B 上蔟後、収繭までを通して25℃目標で保護
C 上蔟後まる6日間を25℃目標で保護し、以後収繭までの短期間を自然温度で保護
収繭後は、各場所の慣習により普通の保護、採種を行ない、交雑種を作った。蚕種は普通に保護した後、12月上-中旬に洗落してバラ種とし、冬期間を通じて毎月1回孵化調査を行なった。
結果の中から群馬県蚕業試験場の成績を示すと第52、53、54表の通りで、産卵状態はA区が最も悪く、B、C両区の間の差は明かでない。
A区は活性化初期の孵化歩合がやや高かったが、最高孵化歩合を示す時期は3区間に殆ど差がなかった。洗落し後を5℃で通したため春蚕催青期には3区とも孵化歩合がかなり低下していたが、特にA区の低下が大きかった。これらの結果は何れも、人工越冬によって試験した第51表の傾向と一致している。
| 蚕品種 | A区 | B区 | C区 | |||||
| 上蔟後 3日間 |
以後収繭 まで |
収繭から 発蛾まで |
上蔟から 収繭まで |
収繭から 発蛾まで |
上蔟後 6日間 |
以後収繭 まで |
収繭から 発蛾まで |
|
| 日124号 支122号(太) |
25.0℃ 25.0 |
19.7℃ 20.0 |
25.7℃ 25.7 |
25.0℃ 25.0 |
25.6℃ 25.6 |
25.0℃ 25.0 |
21.0℃ 21.0 |
25.7℃ 25.6 |
| 蚕品種 | 区別 | 正常蚕卵 蛾 (%) |
少数蚕卵 蛾 (%) |
不受精 卵蛾(%) |
再出卵蛾 (%) |
無蚕卵蛾 (%) |
| 日124号×支122号(太) | A B C |
70 84 81 |
8 9 9 |
13 6 6 |
0 0 0 |
8 0 3 |
| 支122号(太)×日124号 | A B C |
25 84 65 |
15 4 4 |
57 10 28 |
0 0 1 |
2 0 0 |
| 蚕品種 | 区別 | 洗落し 翌日 |
出庫月日 | 春蚕催青 の時 |
|||
| 12月25日 | 1月20日 | 2月20日 | 3月20日 | ||||
| 日124号 × 支122号(太) |
A B C |
2% 0 1 |
62% 40 52 |
95% 95 94 |
95% 93 94 |
96% 94 95 |
66% 67 73 |
| 支122号(太) × 日124号 |
A B C |
3 0 2 |
56 38 40 |
96 92 93 |
95 97 97 |
96 96 97 |
80 92 93 |
ii 人工孵化種 渡辺(1149)は、上蔟から蛾発までの保護温度20℃-30℃の範囲内においては、保護温度の高いほど、有効な孵化歩合の得られる即浸浸漬時間の巾が広いと報告している(第55表)。しかし、足立ら(9,10,11)の現行品種についての試験成績では、品種によっても相違があり、このような明瞭な関係を認め得なかった。
| 浸漬時間 | 保護温度 | |||||
| 30℃ | 25℃ | 20℃ | ||||
| 総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
|
| 30秒 1分 2 3 5 10 12 15 20 25 30 35 40 |
26 90 95 97 93 96 90 93 91 96 68 19 4 |
19 78 84 92 89 91 89 92 90 90 67 17 3 |
40 97 98 98 98 95 98 98 94 75 81 20 0 |
37 96 98 97 98 94 98 97 91 72 70 14 0 |
81 96 97 96 96 97 91 88 79 31 33 12 0 |
74 81 97 96 96 96 86 63 61 20 20 6 0 |
短期冷蔵浸酸についての渡辺の成績は第56表の通りで、即時浸酸の場合と同様、90%以上の孵化歩合の得られる浸酸時間の巾は上蔟後の保護温度の高い場合に広かった。
渡辺は、蚕の化性は上蔟から発蛾までの期間において保護温度の影響をうけるので(1139)、人工孵化の成績も種繭保護温度の高低によって相違するのではないかと考えて、この試験を行なったと述べているが、これらの結果と化性との関係については言及していない。
第55表をみると、90%以上の孵化歩合を示す浸漬時間の巾はなるほど30℃区が最も広いが、最も短い浸酸時間での孵化歩合をみると、保護温度の低いほど高いから、低温区において浸漬時間の巾の狭いのは、塩酸刺戟に対する感受性が強く、浸酸時間が長いと被害を受けて孵化し得ないのであって、低温保護のために越年性が安定して浸酸が効きにくいのではない。上蔟後の保護温度が低いと卵が小さく(Ⅲ3Bb)、卵が小さいと塩酸刺戟に対する感受性の強い(Ⅷ1A)ことがこの原因ではないかとも考えられるが、明かにされていない。
| 浸漬時間 (分・秒) |
保護温度 | |||
| 25℃ | 20℃ | |||
| 総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
総孵化歩合 (%) |
最多2日間 孵化歩合(%) |
|
| 1・0 1・30 2・0 2・30 3・0 5・0 6・0 7・0 8・0 10・0 12・0 15・0 |
93 91 96 96 98 97 95 72 68 2 2 0 |
87 85 92 93 95 90 79 70 67 2 2 0 |
73 95 96 96 96 93 73 50 21 4 1 0 |
63 87 94 94 95 92 71 46 16 2 0 0 |
沓掛(475)は、上蔟から発蛾までを25℃と15℃ とで保護した種繭からの卵の即時浸酸について、第55表と同様な成績を示しているが、上蔟から化蛹までを前期、化蛹から複眼着色までを中期、以後発蛾までを後期として、25℃と15℃とを組合わせて時期別に保護温度を違えると、90%以上の孵化歩合を示す浸漬時間の巾に最も大きく影響するのは中期の温度であると云う。この時期の保護温度が卵の大小や休眠に影響することは既に述べた。
冷蔵浸酸は、産卵後約2日間高温で保護して越年性を進めた卵を冷蔵後に処理するのであるから、即時浸酸の場合と違い、越年性が成績に影響する筈である。第56表において、卵の小さい筈の20℃区の孵化歩合が、最短浸酸時間では最も低く、最高孵化歩合を示す浸酸時間を超えると、他の2回に比べて早く低下しているのは、越年性と卵の大きさとの両者が共に関係していることを示すものであろう。
小野寺(715)が、支110号の同蛾区の熟蚕を3分して、上蔟後、発蛾までをそれぞれ30℃、25℃および20℃に保護したものから採種し、同一母蛾の卵を分割供試して行なった蚕種の抵抗力試験によると、即時浸酸後3日目から40℃、湿度75%に、24時間接触した場合の抵抗力指数(対照区の孵化歩合を100とする)は、30℃区(供試蛾数37蛾)64.8±1.9、25℃区(23蛾)48.5±2.1、20℃区(37蛾)44.2±1.5で、上蔟後の保護温度の高いほど抵抗力が強く、特に30℃区の指数は著しく大きかった。
また、浸酸後3日目の卵を蒸溜水に24時間浸漬した場合には、30℃区(37蛾)70.5±2.5、25℃区(26蛾)44.2±2.9、20℃区(37蛾)36.4±2.8、浸酸後2日目の卵を無水アルコールに50分間浸漬した場合には、30℃区(17蛾)28.8±3.0、25℃区(22蛾)2.7±1.0、20℃区(18蛾)1.1±0.3、4日目の無水アルコール浸酸30分間の成績も同様な傾向を示し、常に保護温度の高い区において抵抗力の大きいことが注目される。
この原因には言及されてていないが、上記渡辺の浸酸試験の場合と共通の原因が関与しているように思われる。
なお小野寺は、点青期における12時間浸水についても同様のことをみている。
4 発蛾調節のための蛹の保護
A 温度管理
発蛾調節のための蛹の保護温度は、蛹期間を短縮または延長させる程度の温度と発育を殆ど停止させる温度(冷蔵)とに区別される。
蛹期間の短縮または延長は、蛹の発育階程に基ずいて計画的に行なうもので、例えば、第38表において、25℃で保護してきた日122号を複眼着色期に28℃に移すと、発蛾までの日数を2日間短縮でき、同じく25℃で保護してきた支115号を複眼黒色期に22℃に移せば、発蛾までの日数を1日間延長することができる。
30℃または15℃を使えば、第41表のように、更に大巾な短縮、延長を行なうことができるが、28℃以上の高温、20℃以下の低温保護は、前節で述べた各種の成績からみて行なうべきではない。なお、蛹の外形に基ずいて温度管理を行なっても、判定の誤差や品種の相違などのために予定通りの調節ができず、次ぎに述べる冷蔵法によって再調整する必要のあることが多い。
発育停止に近い温度による冷蔵法の施行は、その時期、期間および温度について、前法以上に注意をする必要かおる。冷蔵時期とその温度については多くの報告があり、不一致の点も少なくないが、一般に適期とされているのは複眼の着色し初めた化蛹3-5日目頃と発蛾前日であり、温度は2.5℃-7.5℃が最も多く用いられている。
藤本(100)の行なった雌蛹の冷蔵試験によれば、25℃で保護中の蛹を2.5℃に7日間冷蔵し、その後は再び25℃に戻して調べた処、化蛹第1日目および2日目冷蔵のものは冷蔵日数以上に蛹期間が延び、それぞれ18.8±0.04日および18.9±0.04日(無冷蔵は11.4±0.03日)となったが、その後化蛹10日目(11日目に発蛾)までの間に冷蔵したものでは、冷蔵期間以上の発蛾のおくれは起らなかった。発蛾歩合は全体的に低下するが、特に化蛹第1日目および2日目に冷蔵したものにおいて最も低くかった。造卵数は化蛹1-7日目、産卵歩合は1-6日目および9-10日目、不受精卵歩合は1-5日目に冷蔵したものがそれぞれ無冷蔵と大差なく、その他の時期の冷蔵においては総べて無冷蔵より劣った。従って、これらに共通してよかった冷蔵時期は化蛹3日目である。
化蛹3日目の冷蔵につき、2.5℃、5℃、7.5℃、10℃、各7日間の冷蔵を比較すると、発蛾歩合は全般に無冷蔵に劣り、試験した範囲の温度では、その高低と発蛾歩合との間に関係が認められなかった。造卵数は、2.5℃-7.5℃冷蔵、産卵歩合は2.5℃冷蔵、不受精卵歩合は2.5℃-5.0℃冷蔵がそれぞれ無冷蔵と差がなく、共通的によかったのは2.5℃であった。
化蛹3日目冷蔵の場合の2.5℃および5℃冷蔵では、無冷蔵(11.2±0.03日)に比べて冷蔵日数(7日間)だけ蛹期間が長くなったが、7.5℃冷蔵では18.0±0.05日、10℃冷蔵では17.5±0.03日で、何れも、冷蔵日数を加えただけは蛹期間が延びなかった。これは、これらの温度においては、冷蔵中に蛹の発育することを示すものである。藤本の実験結果を綜合すると、蛹期間が10日の場合の化蛹3-5日目(上蔟7日目)に2.5℃に冷蔵すれば、発蛾は多少不斉になるが、その他の点では、最長5日間までは無冷蔵と変りがないと云うことになる。但し、品種は支那二化性S2である。化蛹3日目の蛹の外形は記載していない。
牛込(1127)の成績をみても、化蛹3日目(複眼の着色し初めた時期)の冷蔵が最もよく、発蛾前日がこれに次ぎ、化蛹7日目(生殖外器キチン板が黒色にみえ初める時期)の冷蔵は最も悪い。冷蔵温度は41゚F (5℃)がよかった。
大宮(739)が日122号および支122号(太)の蛹を30℃に3日間、5℃に7日間、または0℃に7日間おいて造卵数を調べた成績では、複眼着色期に処理したものが、腹眼黒色期に処理したものよりも常に造卵数が多かった。産卵歩合には処理時期による差がなかった。
戸谷(1069)によれば、化蛹5日目の蛹(日124号、蛹期間12日)を5℃に10日間冷蔵し、発蛾後解剖して卵管を調べると、卵管下部に卵殻形成の不完全な卵がみられる。化蛹8日目の冷蔵ではこの異常卵の分布する部位が卵管の中部となり、更に冷蔵時期がおそいとこれが卵管の上部に分布するようになる。産卵させてみると、これらの異常卵は早期潰れ卵になるが、冷蔵時期がおそいほどこれの産み出される順位もおくれ、卵管内で認めた異常卵の分布関係とほぼ一致した。異常卵は卵殻に穴のあるもの、または卵殻が内層のみで中、外層を欠くものなどであったが、切片にして調べた結果、異常卵になるのは包卵被膜細胞が完成しかけた時期に冷蔵した卵であることがわかった。第36表について見ても、化蛹5-6日目の蛹の卵管下部は丁度この時期の卵で占められており、蛹令の進むに連れてその部位が上昇する。この実験において(化蛹7日目の冷蔵で潰れ卵が最も多く、3日目の冷蔵では潰れ卵の発現しなかったことは上に述べた冷蔵試験の結果とも一致している。10日間にも亘る冷蔵は実際には行なうべきものではないが、冷蔵は蛹の発育階程を考えて行なう必要のあることを示す好例である。
横山・丸茂(1200)は日122号(細)および支115号を用い、上蔟後25℃、湿度75%目標で保護した種繭を、複眼の着色し初めた頃と発蛾前日とに5℃、湿度80%目標で3日および7日間冷蔵したが、複眼着色期に冷蔵し、出庫時に10℃および20℃各6時間の中間温度を通したものが発蛾歩合、産卵数、孵化歩合、催青死卵、不受精卵などの調査において最もよく、冷蔵3日と7日との間には余り差がなかったと云う。発蛾前日の冷蔵は冷蔵日数の長くなるほど成績が悪かった。
以上の結果を通覧すると、蛹の冷蔵は複眼の着色し初める頃に行なうのが最もよいが、発蛾調節のための冷蔵は初めから予定して行なうものではなく、応急措置であるから、長い期間の冷蔵では劣るとしても、作業上からは発蛾前日に行なうことが多い。
発蛾前日の冷蔵は3日間でも採種成績に影響するが、冷蔵の影響は、裸蛹と繭殻中に収めたものとでも相違する(第57、58表)。
無冷蔵の場合でも、裸蛹にしたものは、切り出し時期の早いほど産卵数が少なく、不受精卵の多いことが古くから云われている(530)。 裸蛹にすると健蛹数が確認できること、発育階程を監視し易いことなどの便宜のある代りに、温湿度などの環境変化の影響を直接に受け易いこと、外傷や病気の感染を受け易いこと、カツオブシムシなどの虫害も多いことなどの欠点を伴なっている。死籠りの多い場合などには切り出し作業に注意しないと、病気を蔓延させることがある。蛹期に膿病ウイルスを接種すると(複眼赤色のとき)、品種によってかなりの遅速はあるが、早いものは82時間(28℃の場合)で発病すると云う(21)。蛹の外皮の最外層は体内水分の発散防止に役立っているから、アルミナの様なもので体表面をこすると体水分の発散が多くなって、正常に発蛾し得ないもののできること、この水分発散防止には脂質の存在が関係しているので、キシロールなどで蛹体面を拭っても水分の発散が多くなり、蛹重の減少すること、などが知られているが(279,283,284)、また外皮の脂質には硬化病に対する抗菌性もあると云うから(410)、切り出した蛹の取扱いは丁寧にする必要かおる。永井(628)は雌蛹の出血が造卵数および産卵数に影響することを報告している。雌蛹の出血は浸酸成績にも影響する(Ⅷ1A、Ⅶ2Aa)。
| 蚕品種 | 試験区 | 発蛾歩合(%) | 産卵状態(蛾の百分率)(%) | ||||||||
| 冷蔵 | 蛹の状態 | ♀ | ♂ | 不産卵 | 少数産卵 | 不受精卵 | 不越年卵 | 累積卵 | その他 | 正常卵 | |
| 日124号 | 無冷蔵 | 切繭中 | 98 | 100 | 4 | 15 | 11 | 0 | 0 | 1 | 67 |
| 3日間 | 切繭中 裸 蛹 鋸屑中 |
92 90 91 |
100 98 99 |
4 2 9 |
20 20 14 |
9 15 11 |
0 0 0 |
1 1 1 |
1 1 1 |
63 60 62 |
|
| 7日間 | 切繭中 裸 蛹 鋸屑中 |
87 79 84 |
100 91 100 |
5 11 7 |
14 20 15 |
14 19 13 |
0 0 0 |
1 0 1 |
1 1 1 |
62 48 61 |
|
| 支124号 | 無冷蔵 | 切繭中 | 93 | 97 | 4 | 8 | 4 | 0 | 1 | 1 | 80 |
| 3日間 | 切繭中 裸 蛹 鋸屑中 |
91 90 96 |
100 95 97 |
5 7 6 |
7 5 8 |
4 5 4 |
0 0 0 |
0 1 1 |
1 1 1 |
81 78 79 |
|
| 7日間 | 切繭中 裸 蛹 鋸屑中 |
83 77 81 |
91 90 91 |
8 7 7 |
8 5 7 |
6 18 14 |
0 0 0 |
2 2 1 |
2 2 1 |
72 63 67 |
|
| 蚕品種 | 試験区 | 正常産卵蛾1蛾当り | 雌1蛾当り | 1g卵数 | |||||
| 冷蔵 | 蛹の状態 | 総産卵数(粒) | 正常卵(%) | 死卵(%) | 不受精卵(%) | 粗卵量(g) | 精選卵量(g) | ||
| 日124号 | 無冷蔵 | 切繭中 | 539 | 95 | 0 | 4 | 0.218 | 0.179 | 1,649 |
| 3日間 | 切繭中 裸 蛹 鋸屑中 |
503 457 526 |
94 96 94 |
0 0 0 |
5 3 5 |
0.198 0.197 0.200 |
0.159 0.167 0.161 |
1,630 1,638 1,615 |
|
| 7日間 | 切繭中 裸 蛹 鋸屑中 |
490 446 472 |
91 88 92 |
0 0 0 |
7 11 7 |
0.201 0.150 0.187 |
0.167 0.111 0.159 |
1,613 1,610 1,613 |
|
| 支124号 | 無冷蔵 | 切繭中 | 526 | 92 | 0 | 7 | 0.254 | 0.237 | 1,773 |
| 3日間 | 切繭中 裸 蛹 鋸屑中 |
509 493 501 |
91 88 91 |
1 2 0 |
6 9 8 |
0.215 0.201 0.205 |
0.202 0.185 0.192 |
1,770 1,780 1,766 |
|
| 7日間 | 切繭中 裸 蛹 鋸屑中 |
477 472 482 |
82 82 82 |
0 0 0 |
16 17 17 |
0.195 0.187 0.189 |
0.172 0.160 0.162 |
1,790 1,771 1,780 |
|
第57、58表の場合は何時頃切り出した蛹であるのか明かでないが、鋸屑に埋めて冷蔵したものの成績が切繭で冷蔵したものと殆ど差異のないのをみると、中間温度を通したものの成績がよかったと云う報告(1200)と合わせ考えて、急激な温度変化によって生理的な障害を蒙ったことが裸蛹の成績を悪くした主な原因と考えられる。蛹に-5℃、24時間または48時間の低温衝撃を与えた場合にも繭層の保護作用が認められると云う。但し、繭腔内温度の測定結果をみると、外温の変化に対する繭層の防護作用は極めて小さい(1193)から、気温ではなく容器の熱伝導が影響しているように思われる。この点を工夫すれば裸蛹冷蔵の被害を軽威することができるかも知れない。
裸蛹で保護するとカツオブシムシの被害を蒙り易いが、新らしい杉の本の鋸屑を日乾したものに蛹を埋めておくと被害が軽減できると云われ、これの手に入り易い地方では実際に用いられている。これは杉鋸屑の匂いがカツオブシムシの忌避剤として作用するのであろうと考えられる。裸蛹を木箔に収容し、杉の精油を加えた籾殻(籾殻100gに対し杉の粗精油0.25-4.00ccの割合に混合)に埋め、蓋をして保護し、発蛾歩合および交尾、産卵を調べたが、蛹に対する杉油の害は認められなかった(875)。杉鋸屑は精油1%内外を含むに過ぎないと考えられるから、カツオブシムシに対する効果は兎も角、蛹の保護材料に用いて害作用はないものと思われる。
B 光線管理
蚕の蛾は、毎朝明るくなると発蛾し初め、数時間内にその日の分は出てしまうのが普通であるが、品種によっては、発蛾がだらついたり、または朝非常に早く発蛾して暴れるために、雄蛾がいたんで採種上の障害になることがある。支115号、支124号などは後者の例であるが、これらの品種においても、光線管理を適当に行なえば発蛾時刻を調節することができる。
第59表は支115号の成績であるが、支124号についても全く同様な結果が得られている。但し、表において8時の発蛾となっているものは27時から8時までの間(8時に近い)の発蛾である。これは、この品種においては、暗中に保護しても12時間までは発蛾を抑えることができず(抑えられるのは10時間以内)、暗中で発蛾し初めるためであって、明暗交替を4時間毎にすると、大部分が明にしたときから発蛾し初める。また、明暗交替の時刻を8時と20時とにとったため、この表では発蛾が8-12時に集中しているが、明暗交替の時刻を希望の時刻、例えば4時と16時とにとれば、発蛾を4-6時に集中させることができる。
明暗交替を始めるのは初発蛾の出る1日前からでよく、収繭後このときまでの明暗は関係がない。明るさは種繭を収容した箔の上面で5lux以上ならば大体よいが、40lux以上あれば申分ない。ただし、暗くしておく間は完全な暗黒が望ましく、5lux程度のうす暗がりでは暗としての効果がない。
蛹の眼、頭頂白色部および腹部に即乾性黒色エナメルを塗ったり、ゴム板で遮蔽したりして試験をしてみると、程度の差はあるが、体表の大部分が光に対して感受性を持っている(434)。しかし、主要な光感受部位は頭胸部にあると推定される(423)。眼と頭頂白色部との何れの光感受性が大きいかはなお不明である(424)。
| 光線管理 | 性別 | 6月13日 | 6月14日 | 6月15日 | 6月16日 | ||||||||||||||||||||
| 4 | 8 | 12 | 16 | 20 | 24 | 4 | 8 | 12 | 16 | 20 | 24 | 4 | 8 | 12 | 16 | 20 | 24 | 4 | 8 | 12 | 16 | 20 | 24 | ||
| 収繭(上蔟7日 目)以後発蛾 終了まで恒明 |
♀ ♂ 計 |
3 1 4 |
5 17 22 |
9 15 24 |
9 6 15 |
8 4 12 |
7 9 16 |
13 12 25 |
21 25 46 |
5 2 7 |
8 6 14 |
3 1 4 |
3 3 |
4 2 6 |
2 2 |
1 1 |
2 2 |
3 3 |
|||||||
| 収繭以後 発蛾終了 まで恒明 |
♀ ♂ 計 |
3 3 |
4 2 6 |
3 8 11 |
4 4 |
2 13 15 |
13 25 38 |
16 11 27 |
12 12 24 |
3 7 10 |
3 3 6 |
5 9 14 |
46 9 55 |
13 13 |
2 2 |
1 1 |
2 1 3 |
1 1 2 |
1 1 |
||||||
| 収繭後恒明、 初発蛾当日 から明暗交替 |
♀ ♂ 計 |
3 2 5 |
11 11 |
3 3 6 |
2 6 8 |
20 20 |
66 58 124 |
24 3 27 |
15 10 25 |
10 10 |
4 1 5 |
||||||||||||||
| 収繭後恒明、 初発蛾1日前 から明暗交替 |
♀ ♂ 計 |
5 31 36 |
8 2 10 |
33 57 90 |
28 5 33 |
1 1 |
1 1 |
19 26 45 |
12 2 14 |
1 1 2 |
|||||||||||||||
| 収繭後恒暗、 初発蛾1日前 から明暗交替 |
♀ ♂ 計 |
2 9 11 |
2 2 |
11 67 78 |
36 1 37 |
1 1 |
1 1 |
42 21 63 |
29 1 30 |
4 3 7 |
2 2 |
||||||||||||||
5 蔟中および蛹の保護環境と各種の障害
A 温湿度による障害
日124号、日126号、日127号などに多発することのある半蛹(半化蛹)は稚蚕28℃、壮蚕22℃で軟葉を給与して飼育すると発生し易いが(Ⅱ2Aa、Ⅱ2Ba)、このような多発環境で飼育したものでも、蔟中の温湿度が低いと半蛹の発生が抑えられ、蔟、繭中の斃蚕も少ないと云う(第60表)。 なお、吐糸終了までを22℃に保護したものを28℃に移すと、全期間22℃保護のものに比べて半蛹の発生がやや多かったから、吐糸終了後の温度も幾分影響するものと考えられる(313)。
| 蚕品種 | 飼育条件 | 蔟中 | 化蛹蚕 歩合(%) |
半化蛹蚕 歩合(%) |
蔟・繭中斃蚕 歩合(%) |
||||
| 温度(℃) | 湿度(%) | 春 | 初秋 | 春 | 初秋 | 春 | 初秋 | ||
| 日124号 | 多発* | 28.0 28.0 25.0 25.3 22.5 22.3 |
80 82 85 80 56 61 |
10 25 72 |
0 14 77 |
35 55 13 |
48 43 8 |
54 18 14 |
41 42 14 |
| 少発** | 28.0 28.0 25.0 25.9 22.4 22.4 |
81 82 86 81 57 58 |
18 25 84 |
68 64 75 |
72 61 6 |
7 7 2 |
9 13 9 |
24 27 22 |
|
| 日126号 | 普通 | 28.0 27.8 28.0 27.8 21.8 22.0 22.4 22.0 |
80 83 55 56 89 90 58 57 |
28 40 55 75 |
53 75 81 84 |
54 50 31 12 |
15 5 7 3 |
18 9 13 13 |
31 19 12 12 |
| 日127号 | 普通 | 28.0 27.8 28.0 27.8 21.8 22.0 22.4 22.0 |
80 83 55 66 89 90 58 67 |
11 33 47 79 |
51 71 66 82 |
58 51 21 11 |
13 5 9 2 |
30 15 31 9 |
36 24 25 16 |
繭の縦作りもこの半蛹の発生を多くし、上蔟後60時間(24.8℃-25.5℃)縦にしておいたものは、その後横にしても矢張り半蛹が多かった(830)。 福田(108,110)の半蛹系統においても、湿度80%のとき、蔟中温度30℃では77.8%、25℃では27.8%、20℃では17.4%と、高温ほど半蛹の発生が多かったが。湿度は65-85%の間では半蛹の発生に影響をおよぼさなかった、95%では増大した。この系統では縦作り繭よりも横作り繭に半蛹が多く、前者47.1%、後者77.6%であった。この半蛹と佐藤らのものとの本質的な異同は明かでないが、福田のものは、脱皮殼を取除いてみると蛹皮が形成されており、また、その下には後に蛾皮が形成されるから、蛹化または成虫化ホルモンの異常によるものではなく、単に、脱皮過程が何らかの原因によって妨げられて生じるものと考えられている。仲野(660)の不吐糸蚕は蔟中温度が25.5℃のときよりも21.5℃のときに却って発生が多いと云う。
日127号、日126号、日124号などには、蔟中温湿度が高いと、ぼか繭、皿繭などの異常繭が発現し、日新にも僅かながらみられるが、日127号には特に多く、23℃-24℃、80%の蔟中環境においても発現することがある(565)。
上蔟以後の保護温度が高いと不受精卵が多くなるが、これがオキシキヌレニンの自働酸化によって着色して、死卵と見誤まられることがある(65,828)。
清水(842)によれば、蛹期間を通して30℃のような高温で保護すると、不受精卵や累積卵が多くなる。また普通には主として卵の裏面にだけ付着している膠着物質(Ⅰ2Ba)が卵の表面にも付着している。これは産卵の機構が異常になるためであろう。この膠着物質は、卵をエオシンの1%水溶液に浸すと赤く染まって容易に観察できる。発蛾前後に5℃に1週間以上冷蔵したものの卵にも同様なことが観察されるが、この場合には包卵被膜をかぶった未完成卵も産み出され、これは産卵後2、3日で潰れ卵になると云う。
三谷(539)は、高温(32℃、80%)と常温(23℃、80%)とに種繭を保護し、その蛾を互に掛け合わせて造卵数、産卵数および不受精卵数を調べ、また雄蛾の貯精嚢および交尾後の雌蛾の交尾嚢、受精嚢の肥大状況、精子の存否、ならびにこれらから取出した精子の運動力(大村法(751))などを調べて、高温保護が雄に悪影響をおよぼすことを確認した。
須貝・木口(870)は、化蛹後の雄蛹を種々な期間33℃で保護して更にこの点を追究し、蛹の後期に比べて前期が高温の影響を受け易いことを報告した。例えば、化蛹直後から23℃
で保護しておいた雄蛹を末期2日間だけ33℃においた場合には、正常雌蛾に交配すると約16%の不受精卵を出すに過ぎないが、末期6日間を33℃
におくと100%の不受精卵を出した(第18図)と云うのであるが、この図は不受精卵歩合の累積曲線であるから、時期別の感受性は明かでないように思われる。精子の量や運動力に異常があり、受精嚢にまで到達し得る精子のないこと、精室から脱出不能の精子が多いこと、脱出しても形態的に異常なものの多いことも観察された。なお無核精子(Ⅰ3Ab)の形成は高温によって著しく阻害されたが、これは無核精子の形成が主として蛹期に入ってから行なわれるためであろうと云う。
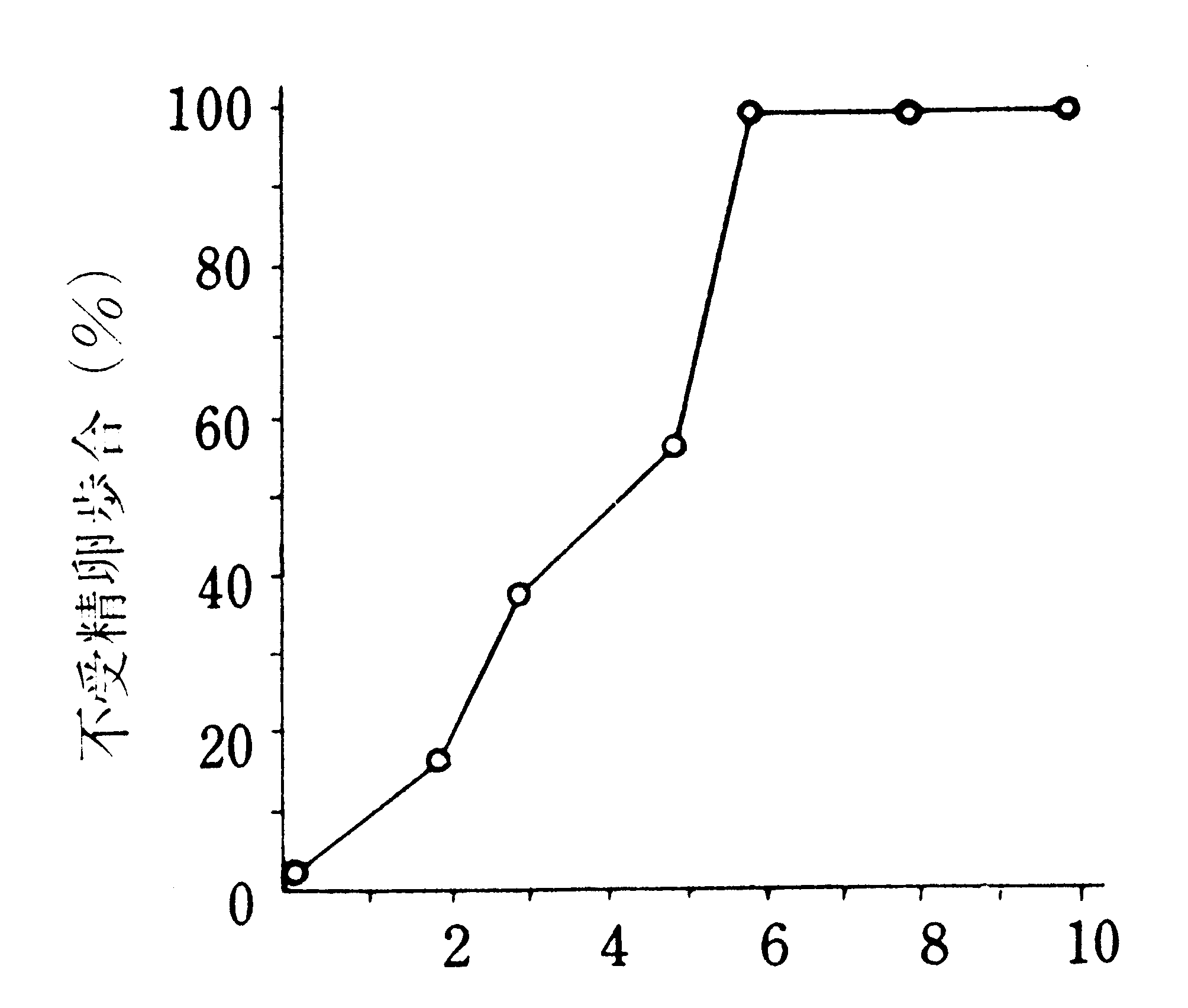
逆算した高温保護日数(日)
第18図 化蛾日から逆算した蛹の高温保護日数と不受精歩合(金色、33℃)(須貝・木口(870))
しかし、その後の研究によると(871)、化蛹後よりも、吐糸から化蛹までの前蛹期の方が高温に対して一層敏感で、雄の生殖減退を起こし易く、正常雌に交配したときの不受精卵歩合は、吐糸後1日間の32℃接触によって12.84%(23℃ の対照区では7.24%)、2日間の接触で29.00%、3日間で72.22%、4日間で89.52%にも達したが、32℃と23℃の接触を12時間々隔で交替すると、8日間続けても不受精卵歩合は対照区と変りがなかった。即ち、高温が244時間以上続くと雄の生殖減退が著しく高まるのである。有核精子の形成が行なわれている幼虫期の高温接触の影響をみるために、4令初期4日間、5令末期3日間の32℃接触も試験したが、雄による不受精卵歩合に影響がなかった。これは、給桑が十分であれば幼虫期の高温による雄の生殖減退が防止できることを示すものであろうと云う。蛹に45℃で8-24時間の高温度衝撃を与えた場合にも精子の運動カが阻害される(701)。
梅谷の陰茎付属筋肉の退化に基ずく不受精蛾の系統(1084,1122)においては、蛹の保護温度が高い(30℃)と異常蛾の発現が多く、しかも上蔟後30℃に保護したもののは5日目頃、20℃に保護したものは6日目頃の1日間だけがこの高温に対する感温期に当ると云う。
種繭を堆積すると内部の温度が高まる。例えば、外温は約18℃から漸次下って10時間後に約15℃になったにも拘らず、3貫500匁の生繭を布袋に詰め、大きさ70×40×30cmの竹龍に入れたものの中央部の温度は、この間に約5℃上昇し、外温との開きが約10℃に達したと云う測定結果もある(844)。外温の高いときはなお更であって、高温と共に炭酸ガス濃度をも高めて障害の原因になるから、繭の輸送および入荷した繭の処置には注意が必要である。
低温による障害については前節において述べたが、滝沢・勝野(1005a)、によれば、蛹期間を13℃で保護した蛾の睾丸は25℃で保護したものの睾丸よりも大きい。これは発育がよいためではなく、13℃保護のものの睾丸には説出し得ない未成熟精子が多く含まれているためであろうと云う。
B 炭酸ガスによる障害
炭酸ガス(濃度98%以上)中に一定時間密閉して死亡率を調べると、化蛹後の蚕は食桑中に比べて抵抗力が強い(第61表)。上蔟後、化蛹までのものは化蛹後のものよりも更に強く、この表と同様な試験の結果では死亡率が0であった。
| 蛹令 | 処理時間 | |||||||||||
| 1時間 | 3時間 | 5時間 | 10時間 | |||||||||
| 日1号 | 支4号 | 欧7号 | 日1号 | 支4号 | 欧7号 | 日1号 | 支4号 | 欧7号 | 日1号 | 支4号 | 欧7号 | |
化蛹1日目 2日目 3日目 4日目 5日目 6日目 7日目 8日目 9日目 10日目 11日目 12日目 |
% 0 10 10 0 0 0 10 0 0 10 10 0 |
% 60 40 20 10 0 0 0 0 0 0 10 - |
% 0 10 0 0 0 0 0 10 0 0 10 0 |
% 20 10 0 10 0 10 0 0 0 10 0 10 |
% 10 40 0 10 40 20 0 0 0 0 0 - |
% 0 0 0 10 0 30 0 0 0 0 0 0 |
% 20 10 0 20 0 0 10 10 0 0 0 10 |
% 30 10 40 10 0 0 0 0 0 10 0 - |
% 0 0 0 0 10 10 10 0 0 0 0 0 |
% 100 20 10 0 0 20 0 20 50 10 0 0 |
% 100 10 40 10 40 20 10 10 10 80 70 - |
% 40 10 0 20 50 20 40 40 20 20 50 0 |
しかし、このような蛹そのものの抵抗性とは別に、種繭の保護中に炭酸ガスの影響を受けたものの産卵には不受精卵が多く、その影響は蛹令の進むに連れて増大する(632)。
諸星(598)は、煉炭で補温して種繭を保護すると、不受精卵が数倍から十数倍にも増加する場合があり、その原因は炭酸ガスの影響と考えられると述べている。
煉炭の場合には、炭酸ガスのほかに亜硫酸ガスの影響も考えられる。また、農家が練炭を用いて不完全な補温をする場合には、管理に注意しないと1日の中の最高、最低の温度差が大きく、高温、低温、あるいは変温の害を受けるおそれがあり、これらは何れも産卵数の減少、不受精卵の増加の原因になる。
Ⅳ 採種
1 雌雄分離
交雑種を作るためには、発蛾までに、何らかの方法によって雌雄を分離しておかなければならない。小規模の試験では、種繭を1個ずつ隔離できるように区劃した容器(分離器)に収容して自由交尾を防ぐ方法も採用できるが、蚕種製造の実用には適しない。従って、雌雄を如何にして分離するかが古くから問題であったことは、直径3分ぐらいの竹で間隙3分ぐらいの簀の子を作り、その上に種繭を一粒ならべにし、新聞紙を覆っておくと、発蛾した雄は暴れ廻るうちに落下するが、雌は簀の子に静止していて落ちるものが少なく、自然に雌雄分離ができ、自由交尾は1割ぐらいに止まる、と云う方法を新案特許として出願したと云うこと(239)からも察知できる。
A 性徴による雌雄鑑別
石渡は、1904年に、雌蚕児の第11環節および第12環節には各1対ずつの小点があるが、雄にはこれがないことを発表した(259-263)。この小点は石渡前腺および後腺と呼ばれているものである(第2、19図、Ⅰ2Ba)。雄には石渡腺はないが、第12環節の前端腹中腺にへロルド腺がある(第10、19図、Ⅰ3Ba)。
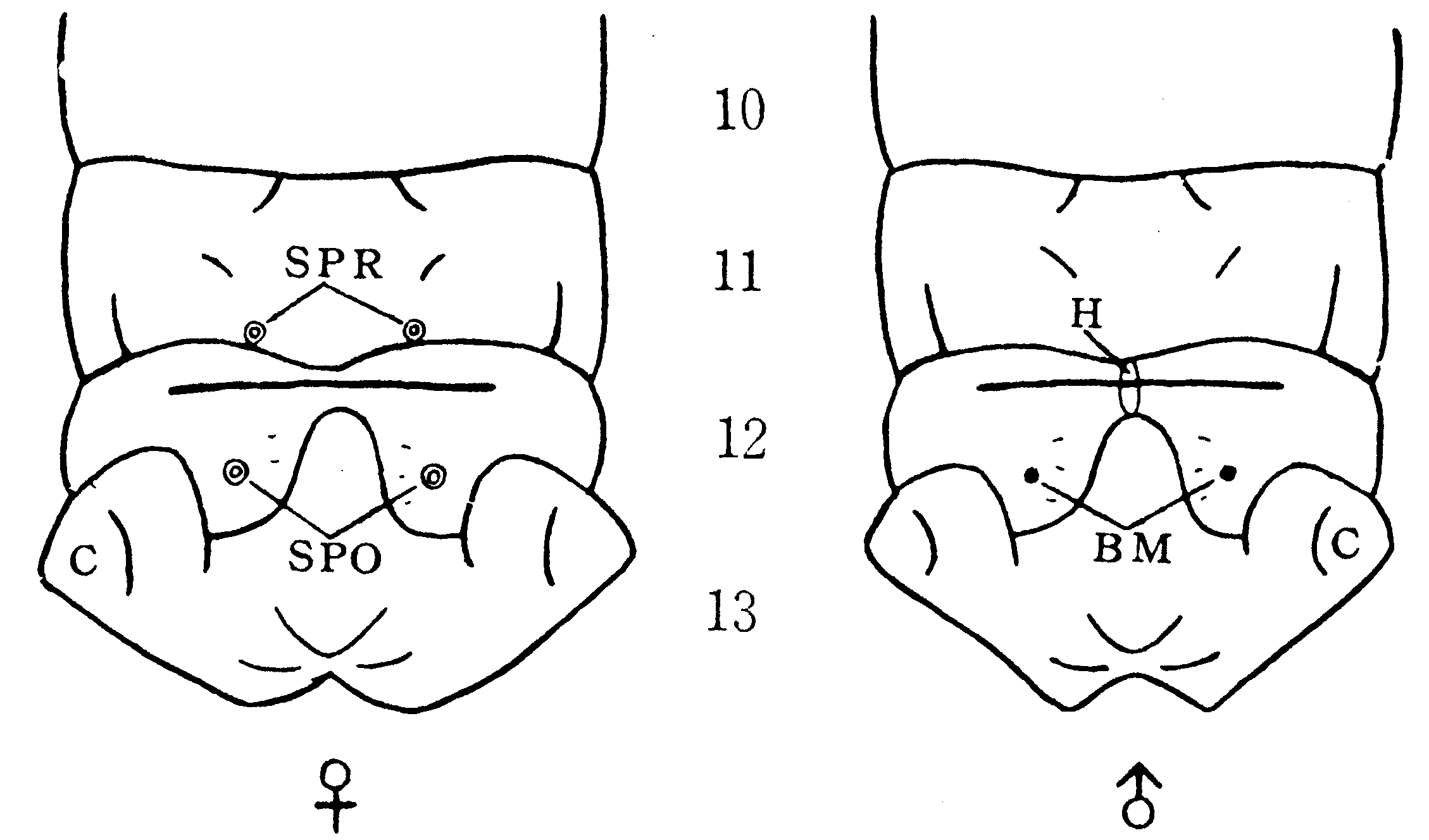
第19図 幼虫の特徴(滝沢・野尻ら)(999)
BM:黒点、C:尾脚、H:ヘロルド腺、SPO:石渡後腺、SPR:石渡前腺、10~13は第10~13環節
この特徴による雌雄鑑別は熟練を要するので、大正11年(1922)に長野県蚕業取締所松本支所において雌雄鑑別手の養成が始められ、鑑別手が蚕種業者に派遣されるようになってから初めて実用的に普及したが、優秀な鑑別手は1日に1万頭以上、1万6千頭も鑑別し、鑑別誤差は0.3%と云われ、わが国の蚕糸業における交雑種の普及を助けた功績は極めて大きい。
石渡腺は5令起蚕において最も明瞭に認められるが、起蚕は取扱いにくく、虫体を損傷し易いため作業に適しない。しかし、5日目には見分けにくくなるので、鑑別作業は5令3日目を中心に行なうのが最もよい。
石渡腺の識別は蚕の品種によっても難易があり、個体によっても不明瞭なものがあるため、加藤(316)はヘロルド腺によって鑑別する方がよいと云い、その適期は5令6日日であるとした。実際には、石渡腺とヘロルド腺とのどちらかと云う訳ではなく、両者を併用して鑑別が行なわれている。イタリアのLombardi(1932)は広東から渡来した大造と思われる品種(White-Jai-ChoおよびGreen-Jai-Cho)においては、石渡後腺に該当する小点だけが1対あって前腺を矢く個体があり、これは雄であったと報告したが、石川(254)は26品種についてこの点を調査し、ヘロルド腺の明瞭な蚕児は、仮令、石渡腺らしいものがあっても総べて雄であり、ヘロルド腺はみえなくても、石渡前腺および後腺のないものも亦雄であることを確認した。この石渡腺らしくみえるものは筋肉の付着点である。第11節の筋肉付着点は石渡も1904年に既に観察して、前腺との区別を図示している。この問題は最近再び取上げられ、滝沢ら(999)が最近の品種について調査を行なったが、大造および輪月では、調査した全部の雄蚕児の第12環節に石渡腺とまぎらわしい黒点があり、支128号および支129号の大多数の雄においてもこれが認められた。この小点は雌の石渡後腺と対応する位置にあるが(第19図)、石渡前腺および後腺が白色の二重環として認められ、周囲との境界が明確であるのに対し、周辺がやや不明瞭な黒点で、石渡後腺よりかやや小さい。その色は品種によって多少異なる。
支115号は石渡腺による雌雄鑑別の困難な品種の一つと云われているが、倉田・四方(462)の調査によれば、この品種に含まれていると云われる雌雄モザイク(Ⅰ2Cb)のために鑑別がむずかしいのではなかった。この場合の鑑別の誤まりは、雄を雌とみたものが966頭中15頭、雌を雄とみたものが1085頭中3頭で、前者の率が高かった。これは上記滝沢らの調査した品種においても同様で、特に、雄の第12環節にまぎらわしい黒点のある支128号および支129号では、この誤まりが多かった。
蛹の雌雄は、幼虫に比べて、識別が容易であるが(第20図)、縦作り繭で尾部の圧縮されたものでは見分けにくいことがある。しかし、蛹では、性徴ばかりでなく、腹部のふくらみ具合によっても雌雄の見当のつくことが多い。

第20図 蛹の雌雄(591)
幼虫鑑別の場合には、蚕をいためないように、取扱いに注意すると共に、作業を始める前に十分に食桑させて、蚕の疲労を少なくするように配慮する。高温の時期には特に注意が肝要である。
蛹体鑑別は、鑑別の誤まりは少ないが、蛹の切り出しに手数がかかり、機械を用いても、両端切りのように能率をあげることがむずかしく、二重繭層のあるものは特に切り出しが困難である。蛹の取扱いについては、裸蛹の冷蔵(Ⅲ4A)について述べたと同様な注意が必要である。
B 限性遺伝の利用による雌雄鑑別
幼虫の体色や斑紋によって雌雄鑑別ができれば便利であろうと云うことは古くから考えられているが、雌雄蚕児の体色や斑絞には特別な違いがないために、普通の蚕の雌雄をそのまま体色や斑紋で鑑別することはできない。
この点について最初に考えられたのは伴性遺伝の利用であった。蚕の性染色体組成は雌がZW、雄がZZで、Wには強い雌性決定因子が含まれているが、普通の形態に関する遺伝子は含まれていない。これに対し、Zには性をきめる作用がない代りに、普通の形態遺伝子が含まれている。それで、目標になる何らかの性質に関して、雌のZには優性遺伝子を含ませ(Z+)、雄のZにはこれに対する劣性遺伝子を持たせておけば(Z-)、第21図のように、これを掛け合わせた子の代の、優性形質を表わす蚕は全部雄、劣性形質を特つ蚕は全部雌であるから、簡単に雌雄鑑別ができる。しかし、伴性遺伝によって雌雄鑑別のできるのはこの1代限りで、これらの子供同志、またはこれを親の代と同じ遺伝子型のものに戻し交雑しても、次ぎの代には、も早鑑別できなくなっている。従って、この方法を利用するためには、毎代、優性の雌と劣性の雄とを準備しておかなければならないので実用に適しない。

第21図 伴性遺伝模式
P:親の代、F:子の代。その他本文参照
限性遺伝の利用と云うのは、これとは反対に、目標になる遺伝子をZ染色体ではなく、W染色体に持たせるものであるが、上に述べたように、普通の蚕はW染色体に形態遺伝子を含んでいないから、突然変異型を利用するのである。
蚕の雌雄鑑別に限性遺伝の利用されるようになったのは、X線照射によって生じたセーブル斑紋蚕と云う突然変異蚕の研究に始まる(1040)。これは第Ⅱ染色体のpの座位に生じた優性突然変異で、暗色蚕と形蚕との中間のような斑紋を表わすが、この蚕の研究中に、セーブル遺伝子がW染色体に転座して、雌に限って常にセーブル斑紋を表わす系統が得られた(1041)。田島はこれを基にして多数の実験を重ね、雌は常に形蚕斑紋、雄は常に姫蚕斑紋を表わす系統を作り、更に実用形質の改良を径て(1042-1045)、最初の限性品種支116号、日117号を育成した。
限性系統利用の原理は、優性遺伝子を持った常染色体の小片をW染色体に転座させ、その性質に関して雌は常に優性を、雄は常に劣性を表わすようにして識別するのであるから(第22図)、斑紋は上記の種類に限らず、虎斑(177)や暗色斑(1048)などの系統も作られているが、卵で雌雄の区別できる系統もある(1047,1049)。

第22図 雌雄鑑別のための限性遺伝模式
白黒の棒:転座常染色体片を持つW染色体(+は優性遺伝子)。
黒の棒:常染色体(-は劣性遺伝子)。縞の棒:Z染色体。
常染色体に関しては雌雄間に差がない。
この限性卵色系統では雌が黒卵(正常卵色)、雄が白卵である。但し、これは普通の雌雄鑑別のためではなく、丈夫で繭質のよい雄だけを飼育しようと云う雄蚕飼育に役立てる目的で育成されたものである。
限性品種は伴性遺伝の利用とは違い、代を重ねてもそのままの特徴が維持されるから、原種の維持は普通の品種と変りがないが、第22図でわかるように、W染色体に過剰な常染色体片が付着していて染色体の平衡が正常でないため、雌が幾分虚弱で、生産性の劣る欠点があった。この付着染色体をできる限り小さくし、欠点を改良する努力が続けられており、一方では雌雄鑑別手が得難くなった事情もあって、今後は利用が拡大するものと思われる。
C 繭重による雌雄分離
雌繭には雄繭よりも重いものが多いので、これを利用して雌雄を分離することが大正年間には行なわれていたが、これは正しい意味の雌雄分離ではなく、重い区分には雌だけが、軽い区分には雄だけが入るように分界点を定めて重量分離を行なうのに過ぎないから、完全に雌雄を分けることは不可能で、中間に雌雄混合の区分ができる。このため一時は殆ど用いられなくなっていたが、最近、鑑別手が得難くなったことと(1168)、各種の商品検査に自働秤りが用いられるようになったこととが結び付いて、再び、新しい種繭重量鑑別機の開発、使用が問題になってきた。
現在発表されている鑑引機には、希望の重量によって、繭を重いもの、中間のもの、軽いものに区分するだけの小型で簡単なものから、計量、分別装置のほかに毛羽焼き機(繭に毛羽が少しでも残っていると、からまり合って自動送繭や分別ができない)を具えた大型装置まで数種あって、大型のものは1時間に4,176粒の繭を処理する能力があり、13日間に1,230kgを処理して、雌423kg、雄430kgを分離したと云うような成績も示されている(837)。しかし、現在では、未だ使用の範囲が狭いため、考案者、製作者側の考え方が先行しており、使用者側の意見や要望が十分に出ていない状態であるから、今後、使用の普及するに連れて一層使い易いものに改善されて行くものと思われる。
繭重による雌雄分離の最大の問題点は、普通の工業生産品の品質管理とは違い、重い繭の多くは雌、軽い繭の多くは雄であるとしても、その中間においては、広い範囲に亘って雌雄が混ざり合っており、その混ざり合いの具合は品種により、蚕期により、更に荷口によって著しく相違することである。このため、分離の良否は機械の性能よりも繭の性状によることが大きい。従って、雌雄を分ける基準重量(分界点、境界点、基準点などと呼ばれている)をどのようにして求めるかがこの方法の実施上最も重要な基本作業である。
繭重による雌雄分離が実際に用いられていた大正時代に広く採用されていた分界点は、
上(♀)の分界点を

下(♂)の分界点を

とするものであったと云うが、新らしい機械の考案と共にこの問題も再び検討されている(682,799,836)。分界点は、現場で数多くの荷ロそれぞれについて求めなければならないものであるから、煩雑な計算を必要としないものが望ましい。
四方(836)は、まず予備分界点によって標本繭の分離を行ない、その結果に基ずいて分界点を補正する次ぎのような便法の例を示している。
予備分界点には上記のもの、即ち

を用いた。但し、この場合には♀--♂-が400mg以上であったが、もし雌雄の差が400mg以下の場合には分母を4以上にするのがよい。
実例として挙げているのは同一分場産の宝鏡についての成績である。まず1荷口につき雌雄各50粒宛の平均を求めると、♀-=1.94g、♂-=1.54gであった。これによって、上式から、♀の予備分界点1.84g、♂の予備分界点1.64gを算出し、これを用いて約300粒の標本繭を実際に機械にかけて分離した後、各区分の繭を切開して調べた処、重い区分の中には雄の混入がなく、雌ばかりであったが、軽い区分の中には雌の1.58gのものが混入しており、中間区分の中で雄の最も重いものは1.76gであった。実用分界点は♀-および♂-に次ぎの補正値を加えれば求められる。
♀の実用分界点を求めるために♀-に加える補正値
=中間区の最も重い♂-♀-=1.76-1.94=-0.18g
♂の実用分界点を求めるために♂-に加える補正値
=軽区中の最も軽い♀-♂-=1.58-1.54=+0.04g
同様の手順によって、他の2荷口につきそれぞれ補正値を求めた処、次ぎの値が得られた。
♀-に加える補正値 -0.21gおよび-0.12g
♂-に加える補正値 +0.07gおよび+0.11g
これら3荷口の補正値を平均すると
♀-に加える平均補正値 -0.17g
♂-に加える平均補正値 +0.07g
となる。この平均補正値は、この分場の、この蚕期の宝鏡に対して、特別に条件が相違しない限りは共通の補正値として使用できるものと考えられる。即ち、他の荷口の繭に対しては、♀-および♂-を求めるだけで、
♀--0.17gおよび♂-+0.07g
をそれぞれ実用分界点として用いることができる。但し、これがあらゆる条件の種繭分離に便法として適用できるが否かは現在の処では明かでないと云う。
分離機を採用して有利か否かは業者それぞれの経営条件によってきまることであるが、技術的には、機械を入れさえすれば、それで100%の雌雄分離がでぎると云うものではないことを理解する必要がある。
D 蛹の体巾による雌雄分離
徳島県蚕業試験場で考案された方法である。体巾による分離と云われているが、蛹の左右の巾ではなく腹背の厚みの相違によって雌雄を分離するのである(287)。蛹の横断面がほぼ楕円形を呈するので、左右の巾を長径、腹背の厚みを短径と呼んでいるが、長径によるよりも短径による方が分別効率(0.5%の誤差で分離される繭の割合)が大きい。岩野らの成績の一部を示せば第62表の通りで、雌雄平均の分別効率は60-70%である。
| 蚕品種 | 性別 | 短径平均 (mm) |
標準偏差 (mm) |
分別点 (mm) |
分別効率 (%) |
調査時期(上蔟後) |
| 春 白 | ♀ ♂ |
9.8 8.8 |
0.30 0.35 |
9.7 9.6 |
58 71 |
春蚕 (11日目) |
| 〃 | ♀ ♂ |
10.0 9.0 |
0.37 0.39 |
10.0 9.1 |
54 59 |
〃 〃 |
| 銀 白 | ♀ ♂ |
9.6 8.3 |
0.41 0.50 |
9.6 8.5 |
47 67 |
〃 (10日目) |
| 〃 | ♀ ♂ |
9.9 8.8 |
0.39 0.30 |
9.5 9.9 |
88 67 |
晩秋蚕 (9日目) |
| 〃 | ♀ ♂ |
10.0 8.9 |
0.39 0.37 |
9.9 9.1 |
66 68 |
〃 〃 |
| 研 光 | ♀ ♂ |
9.4 8.1 |
0.40 0.42 |
9.2 8.4 |
73 74 |
春蚕 (11日目) |
| 〃 | ♀ ♂ |
9.1 8.1 |
0.37 0.39 |
9.1 8.2 |
56 61 |
〃 〃 |
| 豊 年 | ♀ ♂ |
8.9 7.9 |
0.37 0.26 |
8.1 7.9 |
89 61 |
〃 〃 |
| 〃 | ♀ ♂ |
9.1 8.1 |
0.31 0.31 |
8.9 8.3 |
74 74 |
〃 〃 |
装置の原理は次ぎの通りである。周辺が滑かで一端から他端へ向って段階的に細くなっている円棒(1)と、周辺の摩擦を大きくした円棒(2)とを平行にならべ、(1)の細い端が下方になるようにして15°-20°の傾斜角度で取付け、(2)を(1)に対して外側の方向に動力で廻転させる。この上端部に蛹を乗せると、蛹は(2)の廻転に連れて横転しながら2本の円棒に乗って斜面を下り、体巾の小さいものから順次に円棒の間の間隙を通って落下する。(1)が下端ほど細くしてあるのは、斜面の下方へ行くほど間隙を広くするためである。落下した蛹は下に設けた区劃容器に受けるのである。
裸蛹で行なうのであるから、雌雄の混ざった中間区の処理が容易であるが、蛹の取扱いに注意する必要があり(Ⅲ4A)、蛹切り出しの能率のほかに分別効率が60-70%であることも考慮に入れておく必要がある。
E その他の方法
a 物理的方法
比重によって蛹の雌雄分離を行なう方法も考えられた。塩水選を行なうのである。体表に気胞が付着していて誤差の原因になるのが難点とされているが、切り出しの問題もある。
電気抵抗や高周波(189)も試みられたが、思わしい結果が出ていないようである。
蛹の腹部に充満している卵が長波長のX線を強く吸収するならば、繭に入ったままで機械的に雌雄を分けることが可能になるが、卵のX線吸収は予想したほどではなかった(未発表)。
紫外線による繭の螢光色が雌雄によってある程度相違することもあるが、これは品種または系統によって異なり、一般的な雌雄艦別に用いることはできない(188,793)。
b 孵化の早晩と雌雄
1蛾の卵の中で、早く孵化する蟻蚕に雌の多い場合のあることは一般に知られている。この早さの相違は雌雄分離に用い得るほど大きくはないが、原蚕の掃立てに利用すれば雌の割合を多くすることができるから、蚕種製造上若干の実用的な意味を持っている(437)。
室賀(614)は、30分間以内に産んだ日新種の不越年卵10蟻区を温湿度および光線の一定な場所(25℃、75%、マツダ40W昼光色電球から1m)で孵化、させ、その蟻蚕を、各蛾区とも、孵化の順序によって10区に分け、各順位毎にこれを混合して掃立て、4令中期まで飼育して雌雄を調べたが、早く孵化した区には明かに雌が多かった(第63表)。
| 雌雄率 | 孵化順位 | |||||||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | Ⅶ | Ⅷ | Ⅸ | Ⅹ | |
| 雌率(%) 雄率(%) |
66 34 |
56 44 |
53 47 |
48 52 |
45 55 |
50 50 |
51 49 |
40 61 |
47 53 |
40 60 |
| 調査頭数 | 152 | 144 | 148 | 149 | 168 | 160 | 162 | 152 | 169 | 143 |
処が、用いた品種は異なるが、即時浸酸種、人工越冬種(産卵後25℃、15日で7.5℃に50日間冷蔵)および短期冷蔵浸酸種(産卵後25℃、48時間で5℃に冷蔵、15日で出庫、浸酸)においては雌の孵化が雄よりも早いとは云われなかった。また越年種には、雌の孵化の早い場合とその傾向の不明瞭な場合とがあった。これらの結果から、室賀は、胚の発育速度は元来雌が雄より速いのであるが、種々な条件によってこれが不明瞭になり、休眠に入るまでは雌雄の発育速度に差があっても、休眠に入るとこの差がなくなり、活性化する時期には雌雄による違いがないためにこのような結果になるものと考えている。しかし、この場合、それぞれの試験に用いた蚕品種が同一でないので、果してこのような結論をしてよいかどうかは問題である。
これに対して、清水、古和田ら(224,225,438,439,857,858)は、人工越冬種において、雌の孵化が早いばかりでなく、活性化そのものも雄より雌が早いのではないかと考えられること、低温催青によって雌雄の早さが逆転して雄の早くなる品種と変化のない品種とのあること、および品種によって、雌の早く孵化するものと雄の早く孵化するものとがあり、この性質は即時浸酸種、冷蔵浸酸種、越年種の何れにおいても変りがないことなどを報告した。最近の蚕品種(原種)には雌の孵化の早いものが多いとも云われている(845,860)。
永友(647,648)、諸星(597)らは、雌雄による孵化の早晩を、それぞれ晩成遺伝子の作用と化性との関係、および晩成遺伝子の作用とホルモン作用の拮抗との関係によって説明している。
1蛾区のうちで最も早く点青した卵を集めて孵化させ、飼育してみると(日122号および日124号)雌が多い(221)と云う。このことは、点青期までの発育の速さに差のあることを示すもののようであるが、点青後6時間または12時間光線を遮断しておくと、雌雄による孵化の早晩の関係が変化し、明所で雌雄差のない品種においては、暗保護の時期が孵化期に近いほど、雌より雄の性比が早目になり、明所で雌の孵化の早い品種においても、暗保護によって雄の発育が進むため、孵化の雌雄差が短縮する(990)と云うから、雌雄による孵化の早晩の問題には、雌雄による点青期以後の光感受性の違いも関係しているように思われる。
支127号の越年種には白ハゼ卵の多発する性質があるが、白ハゼ卵多発蛾区の生き残った卵から孵化した蚕(外観正常な卵のほか、白ハゼ卵の一部も生き残って孵化する)には雄が多かった(226)。しかし、品種は異なるが春採り越年種を長期間冷蔵しておいたために孵化不良になった蛾区の蚕においては、このような傾向を確認することができなかった(850)。
滝沢・大友(1001)によれば、日124号、支122号(太)の越年種および瑞光、日127号の即時浸酸種において、20%以上の不受精卵を出した蛾回(一部は雌蛾を冷蔵して不受精卵を多発させた)から孵化した蚕は、性比も飼育成績も対照区と変りがなかった。
堀内・清水(222)は孵化の早晩と蟻蚕の体重との関係を調べ、孵化の早いものは体重が小さいが、これは雌雄とは関係がないことをみている。これは、初産卵が晩産卵に比べて大きく(242,408,1142)、大卵は小卵よりも催青日数が長く、それから孵化する蟻蚕の重いこと(Ⅱ2Ac、Ⅲ3Bb)、卵の大小と雌雄とは関係のないこと(242,614,1142)などから当然考えられる結果である。
卵の産み着けられる順序と雌雄との間に関係のないことは限性黒卵(Ⅳ1B)を使って観察した原田・真野(150)の結果、および大沢(773)が3蛾区全部の卵を産み着け順に区別して行なった1頭育の結果からみても明かである。
なお、横山(1195)は、台紙に産み着けられている卵を表面から観察し、孵化前日の胚が体の右側を上面(表面)に向けている卵から雌が多く孵化し、左側を上面に向けているものからは雄が多く孵化すると云っているが、卵内における胚の左右位と雌雄との関係は、古く明石(38)により、また後には石井(250)によって否定されている。
2 発蛾
A 発蛾の行動
a 発蛾および脱繭
川浪(363)は発蛾および脱落の行動を次ぎのように観察している。
発蛾が近付くと、蛹は体表に皺が生じて柔軟となり、脱皮の30-60分前から体を動かし初める。
20-40分前頃から、脚、翅、腹部環節の順序で帯褐乳濁色に変色し、変色が完了すると10-30秒間ぐらい屈伸運動が続き、蛹皮が破れる。蛹皮が最初に裂開する部位は胸部背面正中線が最も多く、胸部と腹部との境の背面がこれに次ぎ、頭部と胸部との境の背面が最も少ない。蛹皮が裂け初めてから脱皮し終わるまでの時間は雄が雌より短かい。
蛹皮から蛾の頭部が出たときに最初の吸胃液の吐出が行なわれる。繭層の一部を切り取って脱落行動を観察すると、まず繭層の吸胃液によって浸潤した部位を第1胸脚でかき分け、頭部をあててその部分を押し、腹部環節を伸縮してこれに力を添える。全部の脚が頭から出ると、脚を繭の外面その他にかけて、体を引き出すような運動を行ない、腹部は伸縮運動を続ける。
脱繭は光の方向、繭のおかれた角度などとは無関係に、常に破風部付近を破って行なわれ、繭の胴部から出るものはない。また明暗や繭のおかれた角度によって脱出の方向を変えるものは極めて少なく、頭部に近い破風部から真直ぐに出るものが大部分であるから、脱繭に際しては走光性を殆ど示さないものと思われる。
吸取紙またはセロファン紙で繭形の袋を作り、脱皮直後の蛾を入れて吸胃液吐出回数を調べたところ、平均して、特N(欧)の雌が35.0回、雄が44.2回、日116号の雌28.1回、雄38.2回、セクザートの雌27.2回、雄36.4回で、何れも雌の吐出回数が雄より少なかったが、平均吐出量は日116号雌201.4mg、雄178.5mg、セクザート雌210.8mg、雄198.6mg、支108号×日115号(新)雌233.4mg、雄221.2mgで、雌が雄よりも良かった。吐出量は微粒子検査用の沈降管内に吐出させて測定した。但し、自然の脱繭よりも長い時問吐出させて測定しているので、吐出回数、吐出量ともに自然の場合よりは多くなっているものと思われる。吸胃液吐出量と蛹重との間には正の相関々係が認められた。
第1回の蛾尿排泄は、脱繭の途中、特に胸部末端から腹部にかけての部分の脱出する際に行なわれることが多い。これは狭い脱出孔を通過するときに直腸嚢が圧迫されるためであろう。次ぎに多いのは脱出後で、繭腔内にあるときに放尿するものは最も少ない。
裸蛹にしておくと吸胃液の吐出量が少ないが、このような蛾と吸胃液を十分に吐出させたものとを比較すると、前者の蛾尿量が多い。尿色は、初めは赤褐色であったが、排出を重ねるに連れて淡色になり、白濁や透明に近い尿を出すものがあった。これは吸胃液不吐出蛾に多く、また透明に近くなった蛾尿はアルカリ性を示した。これらのことは、残存吸胃液が腸を通り、直腸に入って尿と共に排泄されることを示すものと考えられると云う。
塚越(1086)の調査では、吸胃液吐出量は、上記とは反対に、寧ろ雌より雄において多かった。
蛾の下顋(小顎)は発蛾後には萎縮して、感覚器管としては殆ど用をなさないようにみえるが、発蛾前にはその腺細胞の活動がみられ、蛹皮と下顋との間に分泌物を出しているから、発蛾の際の脱皮液分泌組織の1部として役立っているのであろうと考えられている(1082)。
b 脱繭不能蛾
脱繭不能蛾は蛹皮をぬいでも繭から脱出し得ない蛾で、繭内蛾とも呼ばれる。繭層の厚い原種、特に支那種にはこれが多いので、繭の両端切りを行なうのが普通である。日本種は両端切りを行なわない場合が多い。
脱繭不能蛾の発生は蚕品種によって著しく異なるから、遺伝的性質に基ずくことは確かであるが、同一品種においても、そのときどきの条件によって増減がある。
小針・金子(390,391)は、脱繭不能の原因が蚕にあるのか繭にあるのかを知るために種々な実験を行なった。
それによると、脱繭不能蛾は、一般に、雄よりも雌に多く、繭形に関しては、同一品種内では、繭長(L)と繭巾(W)との比率a=L/Wが1に近いほど、即ち球形に近いほど、多かった。また、一般的に云えば雌の脱繭不能が多いのであるが、比較的球形な繭についてみれば雌よりも雄の脱繭不能が多く、長目の繭では雌の脱繭不能が多かった。
繭層を直径0.5cmの円形に打ち技き、その重量によって繭層の厚さを比較すると、縦作り繭においては、蛹の頭部の方向の破風部は尾部の方向の破風部よりも遙かに薄く、横作り繭では両破風部の厚さがほぼ等しかったが、これらの繭の胴部を切り開き、蛹の頭尾を反対にして繭を接ぎ合わせておくと、縦作り繭で蛹の向きを反対にした場合には脱繭不能蛾が多くなるのに対し、横作り繭では繭の向きを変えても脱繭不能蛾は増加しなかった。即ち、繭層の厚さが脱繭不能に関係していることがわかる。
次ぎに、同一品種内で球形に近い繭(a=1.21-1.36)と長形の繭(a=1.48-1.64)とを選び、
1.球形繭の蛹を他の球形繭に入れたものと長形繭の蛹を球形繭に入れたものとの比較
2.球形繭の蛹を長形繭に入れたものと長形繭の蛹を他の長形繭に入れたものとの比較
を雌雄別に、5品種について行なった処、1品種4組(雌雄各2組)、計20組の比較の中で、
球形繭からの蛹に脱繭不能蛾歩合の高かった場合 11例
〃 〃 低かった場合 4例
両種の蛹の脱繭不能蛾歩合に差のなかった場合 5例
で、球形繭の蛹に脱繭不能蛾が幾分多かった。
しかし、この成績を繭層の側からみると、蛹は球形繭のものでも長形繭のものでも、例外なしに、球形繭に入れたものの脱繭不能蛾が長形繭に入れたものよりも多かったから、球形繭に脱繭不能蛾の多いのには、繭そのものの性質の大きく関係していることがわかる。但し、この場合の試験は、雌繭は雌繭、雄繭は雄繭相互の間で蛹を取りかえ、蛹の向きも、前の蛹の向きと同じにして行なった。
一方、同一品種から、淘汰によって脱繭不能蛾歩合の相違する系統を作り、各系統の繭層に一つの系統の蛹を入れてみると、繭層に用いた系統によって脱繭不能蛾歩合が相違し、一系統の繭層に各系統の蛹を入れると、蛹の系統によって脱繭不能蛾歩合が相違した。即ち、脱繭不能の原因は繭の性質と蛾の脱繭能力との両者に関係のあることがわかる。
環境条件としては、5令の軟葉給与が普通桑給与に比べて、発蛾間近かから暗黒にして保護したものは自然の明暗交替にまかせたものに比べて、それぞれ脱繭不能蛾が多かった。また、完全な暗黒ではなく、高さ4cmの木箔に繭を1粒ならべにし、蚕座紙で作った孔紙を2枚覆い、採種室の棚に挿して上下の木箔の間隔を4cmとした程度の光線遮断でも、木箔に繭を並べ、同じ室の中央に1枚ならべにしたものに比べて、脱繭不能蛾が多かった。
なお、近年脱繭不能蛾の多くなったのは、繭層歩合を目標にする品種改良が進んで、繭層が蛾の脱繭能力以上に厚くなったことが一つの原因と考えられるが、また、品種改良の過程において、裸蛹のままあるいは蛹を切繭に入れて発蛾させ採種することが行なわれるため、本来ならば脱繭不能で除外される筈のものからも採種する機会が多くなり、結果として脱繭不能蛾の多い系統を選出する場合があるに違いないと云う。育種専門家の意見として注目される言葉である。
塚越(1086)は、吸胃液吐出量の多い品種は概して脱繭歩合が高いが、例外もあり、球形の繭では吸胃液を分散的に吐出している場合がみられると云っている。
両端切りを行なった繭においても脱繭不能蛾の多いことがある。その原因には種々あると思われるが、繭の切り方によっては、蛾が出るには不十分であり、しかも吸胃液は作用しにくい穴の大きさになる場合のあることや、二重繭層の甚だしい場合には蛾の動きが制約されることなども原因の一部になるものと考えられる。
B 発蛾の早晩
a 発蛾の早晩と産卵
同一品種の同じ掃立て口の蚕について、発蛾の早晩と産卵数との関係を調べると、早く発蛾したものの産卵数は幾分少ない傾向があったと云う報告と(851)、これを追試してみたが、造卵数は、日本種(2品種)においては発蛾日順の早いものに、支那種(2品種)においては発蛾日順の中程のものに多く、良卵、再出卵、不受精卵、死卵、箱内産卵(2日間産卵させた後収蛾箱に入れた)および体内残留卵と発蛾日順との間には、各品種とも、一定の傾向がみられなかったと云う報告(375)とがあるが、全体で100頭内外の蛾数を調べたのでは、初期および後期には日別発蛾数が少ないから、十分な比較はできないように思われる。
小泉ら(433)が別の目的で行なった試験成績の中から、支124号の発蛾第1日と第2日との分を拾い出して集計してみても、1蛾平均で、第1日427.1粒、第2日431.7粒で、差があるとは云われない。但し、これは第1日の11試験区、第2日の12試験区(原著者が冷蔵の悪影響を受けたかも知れないと考えている1区を除き)の各試験区別平均数の平均であって、蛾数は示されていないが、各区の総卵数(24時間内)とその平均産卵数とから逆算、合計すると第1日263蛾、第2日253蛾についての調査と考えられる。
小泉ら(432)は、支124号において、第1日目発蛾と第2日目発蛾との産卵速度を比較し、春蚕期、晩秋蚕期ともに2日目発蛾のものの産卵が速く、大造においても同じ頑向が認められたが、日1号および日124号においては差がなかったと報告している(第64表)。
| 調査 時期 |
発 蛾 日 時 |
時間別積算産卵数歩合(%) | 対1蛾 産卵数(粒) |
||||||
| 1時間 | 2時間 | 3時間 | 4時間 | 5時間 | 6時間 | 7-24時間 | |||
| 6月 | 1日目 2日目 3日目 |
1 23 9 |
8 61 48 |
14 75 58 |
23 84 78 |
30 87 87 |
34 89 91 |
100 99 100 |
414 436 447 |
| 10月 | 1日目 2日目 3日目 |
0 4 11 |
0 24 31 |
0 32 40 |
1 32 45 |
3 37 68 |
7 52 83 |
100 99 99 |
280 277 327 |
この原因には言及されていないが、差のあった品種の発蛾第1日の産卵は非常におそいが、差のなかった日1号および日124号においては、発蛾第1日の産卵が、割愛後1時間でそれぞれ59.1%および41.4%にも達していたから、この遅速は卵の成熱度に関係のある問題のように考えられる。例えば、第2日の発蛾には、第1日目に発蛾すべきもの、あるいは発育のかなり進んだものが、発蛾の日週期や明暗の関係で2日目の朝まで発蛾せずにいたものの混っている可能性があり、このような蛾には、後に述べるように成熟卵が集積していて、早く産卵することも考えられる。これらの点を吟味して再検討する必要がある。第2日目と第3日目との産卵速度には統計的に有意な差がなかった。
b 発蛾の早晩と化性
中間温度(20℃)で暗催青を行ない、不越年卵性蛾の出方を調べると、発蛾日次の早いほど不越年卵性蛾歩合が高い(219,417,566,846-849,851,859)。
この場合、越年卵性蛾と不越年卵性蛾とを比較すると概して前者の産卵数が多い(第65表)。
本書の範囲外のことではあるが、微粒子病蛾は同じ掃立ロの中で早く発蛾したものに多い(735)。
| 系統・化性 | 品種名 | 越年卵性蛾 | 不越年卵性蛾 | 越年卵、不越年卵混産蛾 |
| 日本種 二化性 |
み や ま 志 賀 ふ じ 日 新 春 月 日122号 |
100 (599) 100 (500) 100 (637) 100 (446) 100 (533) 100 (482) |
86 - 94 105 93 - |
96 95 98 112 99 103 |
| 支那種 二化性 |
ひ か る 美 鈴 さ く ら 和 光 宝 鐘 春 白 支115号 |
100 (439) 100 (547) 100 (730) 100 (574) 100 (643) 100 (366) 100 (279) |
128 91 85 89 87 102 194 |
126 98 93 94 98 111 220 |
3 交 尾
A 交尾時間
a 交尾時間と受精
交尾した雄蛾が第1回の射精を始めるのは、初交の雄では5-10分後、射精の終るのは約40分後であるから、初交の交尾時間は1時間で足りることを既に述べたが(Ⅰ3Ba)、これは実際に交尾させた試験結果からみても十分な時間である(2,498,511,863,961)。
1回の射精によって雌の体内に入る精子の数は約10万と算定されているから(Ⅰ3Ba)、精子の数としても不足することはない。
第1回の射精が終って、第2回が始まるまでには60-90分の間があり、この間は、交尾を続けさせておいてもただ連結しているだけで、受精卵を増加させることには役立たない(第66表)。
| 蚕品種 | 交尾時間(分) | |||||||||||
| 5 | 10 | 15 | 20 | 30 | 40 | 50 | 60 | 90 | 120 | 180 | 240 | |
| 日123号 昭 白 長 光 支123号 新 光 |
0% 0 0 0 0 0 |
0% 0 0 0 0 0 |
0% 0 0 0 0 0 |
0% 8 0 97 0 77 |
94% 98 98 91 97 97 |
91% 97 97 95 98 92 |
99% 85 99 96 88 96 |
82% 96 98 93 96 96 |
96% 94 93 97 93 98 |
85% 95 99 90 84 98 |
96% 97 97 93 95 99 |
88% 96 98 94 98 97 |
従って、作業の手順から、3時間以上交尾させておくのは普通のようであるが、再交、三交に使用するための雄蛾の生理上からは、初交の交尾時間は1時間以上に亘らないことが望ましい。交尾を1時間で打切っても、勿論、次代蚕には影響がない(3)。
第1回の射精後直ちに割愛した雄蛾をそのまま直ちに再交させた場合には、2時間は交尾させておく必要がある(498,863)。これは貯精嚢に精子が充満するのに要する時間、射精管下部の分泌物の回復に要する時間などからみても最小限に必要な時間である(Ⅰ3Ba)。再交では初交に比べて射精量が少なく、射精継続時間も短かい。
町田・渡辺(498)によると、24℃と32℃とでは受精に必要な交尾時間に差がなく、15-20分間交尾させるとその産卵中に受精卵が認められたが、12℃では1時間の交尾で初めて受精卵がみられ、7℃では2時間の交尾で漸く受精卵が認められた(日107号)。交尾中には適当な補温の必要なことがわかる。
b 交尾時間と産卵
初交の交尾時間は産卵数からみても1時間で十分である(第67表)。
| 蚕品種 | 交尾時間 (分) | |||||||||||
| 5 | 10 | 15 | 20 | 30 | 40 | 50 | 60 | 90 | 120 | 180 | 240 | |
| 日123号 昭 白 長 光 支123号 新 光 |
24 7 11 10 8 30 |
10 43 13 13 3 30 |
14 15 9 0 4 11 |
14 5 3 88 12 119 |
100 100 100 100 100 100 |
102 77 103 107 116 117 |
111 91 132 97 144 110 |
99 72 111 100 131 116 |
113 93 117 106 135 124 |
76 103 118 87 110 129 |
106 98 121 106 125 117 |
90 102 138 128 142 141 |
しかし、交尾時間を更に長くすると、一見、交尾時間の長いほど産卵数が多いか、または産卵が速いようにみえる現象があらわれる。
この点については沓掛(471)の試験がある。営繭後25-26℃、湿度70-80%で保護した種繭から、朝の4-5時に発蛾した雌蛾をとり、6時に一斉に交尾させ、8時、12時、16時に割愛(交尾時間はそれぞれ2、6、10時間)した試験(A)および同じく4-5時に発蛾した雌蛾を交尾時間は一定(2時間)にし交尾時刻を違え、上と同じ8、12、16時に割愛した試験(B)を行なって産卵速度を比較した(第68表)。
| 割愛 時刻 |
(A)交尾時刻を一定にした場合 | (B)交尾時間を一定にした場合 | ||||||
| 60%内外に 達する時間 |
80%内外に 達する時間 |
60%内外に 達する時間 |
80%内外に 達する時間 |
|||||
| 割愛後 | 発蛾後 | 割愛後 | 発蛾後 | 割愛後 | 発蛾後 | 割愛後 | 発蛾後 | |
| 時 8 12 16 |
時間 8 5 3 |
時間 11 12 14 |
時間 15 10 7 |
時間 18 17 18 |
時間 8 4 2 |
時間 11 11 13 |
時間 14 8 7 |
時間 17 15 18 |
表の(A)をみると、割愛時刻のおそいほど、従って交尾時間の長いほど割愛後の産卵が早くなっている。しかし、(B)をみると、交尾時聞が一定であるのに、割愛時刻のおそいほど割愛後の産卵が早くなっており、また発蛾後の時間で計算すれば、(A)も(B)も殆ど産卵速度に違いがないから、割愛時刻のおそいほど産卵が早いようにみえるのは、卵の成熟度の相違を示すものに外ならないと考えられる(Ⅳ3Ca)。 また、上と同じ時刻に発蛾したものを、交尾時刻を6、10、14時とし、割愛時刻を一定(16時)にすると各区間の産卵速度に殆ど差がなかった。これらの結果からみると、産卵数および産卵速度に関しても、特に長い交尾時間は必要でない。
なお、割愛時刻がおそいと、割愛後、暗くなるまでの時間の短かいこと(Ⅳ5Ab)も産卵を促進する要因の一つになっているものと思われる。
15時に割愛すると、早く産み初めはするが少量の産卵が続き、最も盛んに産卵するのは17-20時頃で、17時に割愛したものと変りがないから、人工孵化などのためには、なるべくおそく割愛して、17-20時に一斉に産んだ卵だけを使用するのがよい(1128)と云うような意見もあるが、これも、割受がおくれれぼその間に卵の成熟が進み、夕方暗くなるのと相俟って、集中産卵の条件が成立するために外ならない。
B 雄蛾の使用回数
同じ雄蛾を何回交尾させることができるかについては、蚕品種や雄蛾の取扱いによって相違するばかりでなく、何日間にも亘って試験を続ける場合には、相手に使う雌蛾の条件が同じではなくなるから、結果の討検に注意する必要がある。
小田中(699)の成績の一部を第69、70表に示す。
| 掛け合わせ | 交尾 回次 |
交尾時間 | 受精卵 歩合(%) |
死卵歩合 (%) |
不受精卵 歩合(%) |
有効雄蛾* 歩合(%) |
| 分離白1号 × 支106号 |
1 2 3 4 5 6 |
1時間半 〃 2時間半 〃 3時間 3時間半 |
95 92 87 90 82 65 |
0.5 0.3 0.6 0.2 0.9 1.7 |
4.4 6.8 11.7 9.5 16.9 32.6 |
100 100 96 96 89 89 |
| 支106号 × 分離白1号 |
1 2 3 4 5 6 |
1時間半 〃 2時間半 〃 3時間 3時間半 |
94 92 91 89 94 94 |
0.9 1.2 0.9 1.0 1.0 0.7 |
4.5 6.4 7.1 9.5 4.0 4.9 |
100 100 100 100 89 89 |
| 掛け合わせ | 交尾回次 | 交尾時間 | 受精卵 歩合(%) |
死卵歩合 (%) |
不受精卵n 歩合(%) |
有効雄蛾 歩合(%) |
|
| 分離白1号 × 支106号 |
1 2 3 4 5 6 |
第1日 〃 第2日 〃 第3日 〃 |
1時間半 2時間 1時間半 2時間 2時間 3時間 |
95 94 85 61 83 52 |
0.7 3.3* 0 0.4 0 0.1 |
3.9 4.9 14.1 38.2 16.7 47.2 |
100 92 92 92 85 75 |
| 支106号 × 分離白1号 |
1 2 3 4 5 6 |
第1日 〃 第2日 〃 第3日 〃 |
1時間半 2時間 1時間半 2時間 2時間 3時間 |
95 95 94 93 90 86 |
0.3 0.2 0.9 0.5 0.9 1.4 |
4.3 4.3 4.5 6.2 9.0 11.6 |
100 100 96 85 82 82 |
この試験においては、使用後の雄は、その都度直ちに13℃で抑制した。抑制のための容器は綴1尺5寸、横2尺1寸、高さ1尺5寸、周囲は金網張りで、中に20個の小框を吊した。蛾はこの小框に張った布にとまり、抑制中殆ど動かなかった。
交尾、産卵中の温湿度は24℃、75%。産卵数調査は産卵後30日で行なった。この成績をみると、支106号と分離白1号とで程度に相違はあるが、1日1回交尾、2回交尾ともに、第3回次から目立って不受精卵が増加している (Ⅳ4B)。
なお小田中は卵の1,000粒重量の違いおも見出そうと努めているが、卵の大小は雌によってきまるものであるから、これは差のない筈のものである。
C 交尾中の管理
a 温度
上蔟後、温度26℃内外、温度70-80%で保護した種繭から朝の2-6時の間に発蛾した雌蛾をとり、6時から4時間、30℃、25℃、20℃および15℃で交尾させ、10時に割愛、25℃で産卵させて産卵速度を調査した沓掛(471)の成績を第71表に示す。
| 産卵時間 の区分 |
交尾中30℃区 | 交尾中25℃区 | 交尾中20℃区 | 交尾中15℃区 | |||||
| 時間別 産卵数(粒) |
累加産卵 歩合(%) |
時間別 産卵数(粒) |
累加産卵 歩合(%) |
時間別 産卵数(粒) |
累加産卵 歩合(%) |
時間別 産卵数(粒) |
累加産卵 歩合(%) |
||
| 6月28日 6月29日 |
10-11時 11-12 12-13 13-14 14-15 15-16 16-17 17-18 18-19 19-20 20-21 21-22 22-23 23-24 0-1 1-2 2-3 3-4 |
135 163 73 20 27 10 10 12 3 4 6 2 3 1 4 2 1 4 |
24 53 66 70 75 77 79 81 81 82 83 84 84 85 85 86 86 87 |
141 176 79 12 23 20 23 11 8 3 9 5 3 3 1 1 2 4 |
24 55 69 72 76 79 83 86 87 88 89 90 91 91 92 92 92 93 |
35 59 148 77 64 27 65 51 18 22 13 9 3 4 3 1 3 10 |
5 13 35 46 56 60 69 77 80 83 85 86 87 87 88 88 89 90 |
12 7 78 98 42 12 43 75 48 22 41 33 6 16 2 4 2 8 |
2 3 16 31 38 40 48 60 68 72 78 84 85 87 88 89 89 90 |
| *その後の産卵数 | 71 | 36 | 65 | 56 | |||||
| 総産卵数 | 558 | 569 | 684 | 613 | |||||
| 発蛾後 割愛までの 保護温度 |
保護時間 | |||||||
| 4時間 | 12時間 | |||||||
| 産卵数歩合 | ||||||||
| 50% | 60% | 70% | 80% | 50% | 60% | 70% | 80% | |
| ℃ 30 25 20 15 |
時間 1-2 1-2 4-5 6-7 |
時間 2-3 2-3 5-6 7-8 |
時間 3-4 3-4 6-7 9-10 |
時間 7-8 6-7 8-9 11-12 |
時間 1-2 1-2 2-3 2-3 |
時間 1-2 1-2 2-3 2-3 |
時間 2-3 2-3 4-5 5-6 |
時間 4-5 3-4 9-10 7-8 |
沓掛は、これと同時に、それぞれの温度に移した蛾を直ちに交尾させず、4時間おいてから交尾させたもの(各温度に保護した時間は交尾中4時間をも含めて8時間)および8時間おいて交尾させたもの(各温度に12時間)についても同様を調査を行なった。これらの結果を綜合すると、第72表のように、産卵歩合が一定値に達するまでの時間は、発蛾後、割愛までの保護温度30℃と25℃との間には殆ど差がないが、20℃以下では、温度の低いほど目立って良くなる。しかし、保護時間が良くなるに連れて各温度区間の産卵速度の差は小さくなる。これは、温度によって産卵速度に差の生じるのは卵の成熟速度が温度によって異なるためで、保護時間が良くなれば各区の成熟卵数の差が次第に縮まることを示すものと考えられるが、これらの温度に4、8、12時間保護した蛾を解剖して、卵管柄に下降している成熟卵数を調べた結果は(第73表)、この予想の通りであった。
短時間内に一斉に産卵させることは蚕種の保護取扱い、特に人工孵化成績を向上させるために重要であるが、これには、産卵中ばかりでなく発蛾から交尾中の環境にも注意し、25℃目標の温度で保護する必要のあることがわかる。湿度は70-80%がよい。
| 発蛾後の 保護温度 |
成熟卵数(10蛾平均) | |||
| 発蛾直後 | 4時間後 | 8時間後 | 12時間後 | |
| ℃ 30 25 20 10 |
粒 445 |
粒 537 556 532 441 |
粒 588 618 579 486 |
粒 621 700 638 560 |
b 光線
交尾中を暗くしておくと、明るくしておくよりも割愛後の産卵が幾分早いと云う成績がある(第74表)。
| 区別 | 交尾中の 照度 |
産卵中の 照度 |
割愛後の時間別産卵数歩合(%) | 24時間の 総産卵数(粒) |
供試 蛾数(蛾) |
||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 8-24 時間 |
|||||
| A | 500lux | 初め2時間は500lux、 以後は暗 |
0 | 1 | 37 | 9 | 8 | 9 | 7 | 8 | 21 | 10,891 | 27 |
| E | 暗 | 0 | 0 | 46 | 4 | 8 | 5 | 8 | 4 | 24 | 11,637 | 26 | |
| B | 500lux | 恒 暗 | 13 | 16 | 14 | 10 | 5 | 6 | 5 | 6 | 25 | 11,825 | 25 |
| F | 暗 | 24 | 14 | 15 | 8 | 6 | 6 | 4 | 2 | 21 | 12,546 | 26 | |
この試験のように500luxと云うような明るい場所で交尾させておくことは実際にはないであろうが、後(Ⅳ5Ab)に述べるように、産卵中の雌蛾に対しては30luxと600luxとが同様な効果をあらわすことを考えると、交尾中を暗黒または暗黒に近い程度にしておく方が割愛後の産卵は早いと思われる。但し、暗くしておくと、離交した場合に雌が直ちに産卵を始めるおそれがあるから、離交し易い品種においては、交尾中を明るくして、離交した雌の産卵を抑制する方がよいと云う。
勝野(321)は、照度0.2、30および300luxの場所で交尾させた雌蛾を割愛後解剖して、交尾嚢および受精嚢内の精子数をかぞえたが、300lux区においては、他の2区に比べて交尾嚢内における“ちぢれ精子”(Ⅰ3Ab)数の比率の高いことを認め、照度の高い場合には“ちぢれ精子”の受精嚢への移行率が低いのであろうと考えている。
交尾中の環境については、その他、60-80フォーンの騒音(録音で聞かせた)は、30-40フォーンに比べて交尾不能蛾を増加させ、不受精卵歩合および残留卵歩合を高めたと云う報告がある。
4 蛾の抑制
A 雌蛾の抑制
発蛾当日の未交尾雌蛾を2.5℃、5℃、7.5℃および10℃に冷蔵し、出庫後、発蛾当日の雄蛾を交配して産卵を調査した藤本(99)の成績を第75表に示す。
| 冷蔵温度 (℃) |
冷蔵日数 (日) |
造卵数 (粒) |
産卵数 歩合(%) |
不受精卵 歩合(%) |
冷蔵中の産 卵数歩合(%) |
供試蛾数 (蛾) |
| 2.5 | 2 4 6 8 |
554 553 562 562 |
83 87 87 82 |
6 8 12 11 |
0 0 0 0 |
93 89 76 80 |
| 5 | 2 4 6 8 |
550 568 569 572 |
85 89 89 83 |
5 6 9 9 |
0 0.1 0.3 1.3 |
88 88 85 83 |
| 7.5 | 2 4 6 8 |
548 566 558 546 |
85 85 74 69 |
6 9 10 15 |
0.3 9.9 14.8 21.5 |
89 86 82 82 |
| 10 | 2 4 6 8 |
558 571 561 558 |
86 78 60 45 |
6 10 16 24 |
3.6 13.3 30.1 43.0 |
81 88 86 79 |
| 無 冷 蔵 | 553 | 86 | 3 | - | 97 | |
これはS2と云う品種(支那二化性)についての成績であるが、同時に行なったN2(日本二化性)についての試験結果も同様で、最も安全な冷蔵温度は5℃であった。
冷蔵巾の産卵は品種によって著しく相違する。発蛾後5時間以内の未交尾雌蛾(支那二化、支那一化、日本二化、日本一化、欧州一化の各2品種あて)10日間冷蔵して調査した結果によれば、2.5℃においては何れの品種も産卵しなかった。5℃においては支那種(一化性1品種を除き)が5-6日目から産卵し初めたが、日本種および欧州種は産卵しなかった。7.5℃においては、支那種は2-4日目から産卵を始め、一化性よりも二化性の産卵が遙かに多かった。日本種は、早いもの(N2)は5日目、おそいものは8日目から産卵を始め、10日間の産卵数歩合は支那1化性と大差がなかったが、欧州種は産卵しなかった。
10℃においては、支那種は1-2日目、日本種は3-5日目から産卵を始め、欧州種のうち豊黄は10日目に僅かに産卵したが、欧20号は全然産卵しなかった。冷蔵中の産卵は、冷蔵前の高温保護時間の長い蛾ほど多く、交尾したものは未交尾蛾に比べて遙かに多い。
未交尾蛾を1日間5℃に冷蔵したものと、この間を24℃においたものとの交尾後の産卵を比較すると(第76表)、S2においては、24℃においたものの方が産卵数歩合が高かった。これは24℃の方が卵の成熟が進むためであろうが、品種によって相違のあることがわかる。この場合には24℃においたものの方がよかったようにみえるが、交尾前に産卵してしまう場合もあるから、一般に用いることはできない。
| 蚕品種 | 試験区 (℃) |
供試蛾 数(蛾) |
交尾後における | 交尾前産卵 数歩合(%) |
|
| 産卵数歩合(%) | 不受精卵歩合(%) | ||||
| S2 (支那二化) |
5 24 |
96 93 |
75 81 |
6 5 |
0 2 |
| N2 (日本二化) |
5 24 |
87 79 |
67 66 |
9 13 |
0 3 |
なお藤本は、8日間以内の冷蔵では、冷蔵と無冷蔵との間に造卵数の差はないと云っているが、第75表において、4日間以上冷蔵したものの造卵数は無冷蔵のものに比べて僅かながら常に多い。これはN2についての成績をみても同じ傾向であるから、意味のある差と考えるべきで、冷蔵中に卵の成熟が進むのであろうが、冷蔵中の産卵の全然なかった2.5℃のような低温において果して卵の成熟が進むか否かは問題である。しかし、蛾の冷蔵は、後に述べるように短時間でも産卵を促進し、また長く冷蔵すると包卵被膜をかぶったままの未完成卵まで産み出されると云うから(Ⅲ5A)、低温刺戟を受けた卵は出庫後の成熟が進んだり、あるいは未成熟のまま産み出されることがあるのかも知れない。
雌蛾を冷蔵すると、以上のように、その温度および期間によって、生理的障害、卵の成熟、および産卵に対する刺戟の三つの影響が組合わさって結果にあらわれる。従来、短期間の抑制には5℃よりも高めの温度の用いられることが多いのは、抑制中に産卵しない限り、温度の高い方が産卵成績のよい場合のあるためと考えられるが、発蛾後、時間のたった雌蛾は、7.5℃以上では抑制中に産卵を始めることが多いから、抑制の時期および温度に注意する必要がある。産卵の早い品種においてはこの注意が特に大切である。
20℃前後の温度での蛾の生存日数は、雌雄ともに概して欧州種が日本種および支那種より長く、一化性は二化性よりも、雌は雄よりも長い。
B 雄蛾の抑制
雄蛾は雌蛾よりも長期間の抑制を必要とする場合が多い。
雄蛾は動きが激しいから、体力の消耗を防ぐことが抑制の眼目になるが、同時に、抑制しながら交尾を繰返えす場合には、次ぎの交尾までの間に射精に必要な内的条件の回復(Ⅰ3Ba)を進める必要がある。抑制温度とこの内的条件の回復との関係についての調査は行なわれていないようである。
種々な点から考えて、安全な抑制温度は5℃-7.5℃であるが、短期間の場合には蛾の静止ししている範囲内で、これより高い温度を用いてもよい。
13℃での抑制は既に示した(第69、70表)。 5℃での成績を第77表に示す。
三交の成績が目立って低下していることは13℃抑制の場合と同様の傾向である。
| 蚕品種 | 交尾回次 | 供試蛾数 (蛾) |
交尾歩合 (%) |
正常産卵 蛾歩合(%) |
不受精卵 蛾歩合(%) |
| 支126号×日126号 | 初 交 再 交 三 交 |
91 91 91 |
100 100 99 |
86 76 83 |
1 7 6 |
| 支125号×日502号 | 初 交 再 交 三 交 |
100 100 100 |
100 96 96 |
73 76 61 |
2 2 14 |
| 日502号×支125号 | 初 交 再 交 三 交 |
100 100 100 |
100 98 98 |
83 76 67 |
8 20 24 |
| 日126号×支126号 | 初 交 再 交 三 交 |
100 100 100 |
100 99 99 |
88 89 85 |
6 7 11 |
5 産卵
A 産卵中の管理
a 温度
上蔟後、温度26℃内外、湿度70-80%で保護した種繭から朝の2-6時に発蛾した雌蛾をとり、直ちに25℃で交尾させ、4時間後の10時に割愛して25℃、20℃、15℃の3区に分け、光線を遮断して産卵させた成績を第78表に示す(471)。沓掛は、これと同様にして8時間交尾させる実験をも行なっているが、この二つの実験結果を綜合すると(第79表)、30℃区と25℃区との産卵速度には差がなく、ともに20℃区および15℃区より速いが、その差は交尾時間の長くなるに連れて小さくなっている。これは既に縷々述べたように、卵の成熟が進んで各区の成熟卵数の差が縮小するためと考えられる。
産卵中の保護温度は胚発生の面からも考えなければならない。産み出された蚕卵内では、後に述べるように(Ⅵ2A)、直ちに成熟分裂、受精、および核分裂が引続いて進行するから、産卵中の保護には母体の保護と共に蚕種保護の初期が重なり合っている。
| 産卵時間 の区分 |
産卵中25℃区 | 産卵中20℃区 | 産卵中15℃区 | ||||
| 時間別 産卵数(粒) |
累加産卵 歩合(%) |
時間別 産卵数(粒) |
累加産卵 歩合(%) |
時間別 産卵数(粒) |
累加産卵 歩合(%) |
||
| 6月28日 6月29日 |
10-11時 11-12 12-13 13-14 14-15 15-16 16-17 17-18 18-19 19-20 20-21 21-22 22-23 23-24 0-1 1-2 |
141 142 69 126 28 16 12 8 7 5 9 6 2 1 0 5 |
22 44 55 75 79 82 84 85 86 87 89 90 90 90 90 91 |
102 138 117 44 34 37 17 20 12 6 7 2 3 4 3 7 |
15 37 55 62 67 73 76 79 81 82 83 83 84 85 85 86 |
68 113 89 99 49 45 9 11 13 14 10 3 2 4 1 6 |
10 28 42 58 66 73 75 77 79 81 83 83 84 84 85 86 |
| *その後の産卵数 | 52 | 86 | 87 | ||||
| 総産卵数 | 637 | 646 | 631 | ||||
| 産卵中 の温度 |
交尾時間 | |||||||
| 4時間 | 8時間 | |||||||
| 産卵数歩合 | ||||||||
| 50% | 60% | 70% | 80% | 50% | 60% | 70% | 80% | |
| ℃ 30 25 20 15 |
時間 - 2-3 2-3 3-4 |
時間 - 3-4 3-4 4-5 |
時間 - 3-4 5-6 5-6 |
時間 - 5-6 8-9 9-10 |
時間 0-1 1-2 1-2 1-2 |
時間 1-2 1-2 1-2 2-3 |
時間 2-3 2-3 1-2 3-4 |
時間 4-5 4-5 3-4 4-5 |
| 蚕品種 | 供試 蛾数(蛾) |
第1次産卵 中の温度(℃) |
第1次産卵(2-18時) | 第2次産卵(18時-翌朝9時) | ||||
| 正常卵 (%) |
不発生卵 (%) |
不受精卵 (%) |
正常卵 (%) |
不発生卵 (%) |
不受精卵* (%) |
|||
| N6形 | 10 7 10 11 10 |
28 30 32 34 **交尾前34 |
91 64 46 14 - |
6 18 40 70 - |
2 17 13 15 - |
82 85 93 94 90 |
8 6 2 3 5 |
9 7 3 1 4 |
| C18 | 9 8 10 10 8 10 |
25 28 30 32 34 **交尾前34 |
96 98 97 32 7 - |
2 1 2 6 43 - |
0 0 0 61 49 - |
89 95 93 98 98 98 |
8 3 6 1 0 1 |
2 0 0 0 0 0 |
受精を正常に完了させることは蚕種保護にとっては何よりも大切である。上の例においては、産卵速度に関する限り、30℃と25℃との間に優劣が殆どなかったが、蚕種保護、特に正常な受精と云う立場からみればは30℃は受精の障害を起こし易く、極めて危険な温度である(第80表)。
この表の不発生卵は、産卵後暫くは普通の不受精卵と区別がつかないが、1週間ほどで徐々に着色し初め、2週間後には濃淡種々な褐色卵になり、漿膜を形成していたものもあるが、調査のときには全部淡褐色の潰れ卵になっていたと云うから、不受精卵の着色(Ⅲ5A)、不完全な単為発生、受精後間もない死卵などを含むものと考えられる。
佐藤・目崎(819)は産卵後78゚F(20℃)で20-80分の卵を104゚F(40℃)に1時間接触せしめた場合に生ずる不受精卵や死卵の増加を細胞学的に研究し、成熟分裂の異常、卵核の行動の異常および崩壊、精核の行動の異常および退化などを観察している。卵核は精核よりも高温に対する抵抗力が弱い。受精した核が分裂の途中で死滅すると外観的には不受精卵と区別できない卵になる。漿膜が生じてから後に死卵になるものも多い。またポリプロイド核が生じ、これが発生の過程で死滅して不着色卵または着色死卵になる。
野尻の場合にはこれよりも接触温度は低いが、時期的に一致しており、受精の異常に基ずくものと考えられる。
河野(449)は、春蚕期に、産卵後24℃で5時間目の卵と10時間目の卵とを28℃および32℃(湿度75-80%)に移して10日間おき、以後は普通のものと同じ扱いをした処、翌春の催青において催青死卵の多発するのを認め、解剖してみると腹面の環節が重複している畸形胚であった。但し、これはある特定の品種に限って起こると云う。
交尾後の雌蛾を短時間低温にあわせると、常温に戻してからの産卵が促進されるが、その促進効果は雌蛾の状態によって異なり、暗くしただけで直ちに多数の産卵をするような蛾を低温処理しても、それ以上特に産卵の早まることはない(433)。従って、その効果は、主として、集積している成熟卵の産卵促進と考えられるものであるから、成熟卵の集積の少ない蛾に低温処理を施しても、短時間内には効果を期待することがむずかしいものと思われる。
産卵促進のための低温は0℃-15℃の範囲内では効果に違いがないが、5℃-10℃
で30分間ぐらいの処理をするのが適当であろうと云う。処理中の明暗は効果に影響がなかった。鈴木(893)は2.5℃、5℃、10℃の各30分間処理を比較して、5℃が産卵促進に最も効果があったと云っている。
低温処理による産卵促進効果は、上記のように、雌蛾の状態によって異なるが、小泉ら(433)の成績中、最も効果の大きかった例を示すと、割愛後を暗くしておいただけの対照区では産卵数歩合が30%に達するのに5時間を要したのに対し、5℃で2時間処理した後、暗所で産卵させたものは1-2時間で30%の産卵数歩合を示した(支124号)。
b 光線
蚕の産卵は明所におけるよりも暗所において速いが(471)、その明暗の区別は絶対的なものではなく、例えば、600luxと30luxとの照明を1時間ずつで交替すると、600luxのときに産卵が抑えられ30luxのときには促進されるが、30luxと3luxとを同様に交替すると、30luxが前の場合の600luxと同様に産卵を抑え、3luxが産卵を促進する(420)。 3luxと0luxとの交替では、3luxは実際上殆ど0luxに等しい作用をするが、交替を長時間続けると僅かに0luxのときよりは産卵の少ない傾向を現わす。明暗に対するこの反応には、蚕の品種によってかなり相違がある(286,420)。伊藤らの成績の中から数例を示す(第81表)。
| 蚕品種 | 産卵中 | 供試 蛾数 |
割愛後産卵数歩合が一定値に達するまでの時間 | |||
| 温度 | 明暗 | 産卵数歩合30% | 産卵数歩合50% | 産卵数歩合70% | ||
| 日 4 号 | ℃ 24.3 |
明 暗 |
蛾 12 16 |
時間 分 4 ・31 1 ・30 |
時間 分 6 ・18 2 ・32 |
時間 分 8 ・20 3 ・35 |
| 分離白1号 |
明 暗 |
22 24 |
4 ・11 2 ・14 |
5 ・40 3 ・46 |
7 ・38 5 ・47 |
|
| 日106号 |
明 暗 |
24 23 |
4 .17 1 ・37 |
6 ・22 3 ・01 |
8 ・32 5 ・23 |
|
| 日110号 |
明 暗 |
20 19 |
3 ・37 0 ・56 |
4 ・46 1 ・34 |
5 ・50 3 ・02 |
|
| 支101号 |
明 暗 |
22 27 |
7 ・24 0 ・47 |
8 ・55 1 ・18 |
10以上 1 ・49 |
|
| 支103号 |
明 暗 |
15 21 |
2 ・34 0 ・44 |
5 ・46 1 ・13 |
7 ・40 1 ・43 |
|
| 欧 5 号 |
明 暗 |
7 8 |
4 ・12 0 ・41 |
5 ・10 1 ・34 |
5 ・49 5 ・12 |
|
| 欧19号 |
明 暗 |
6 9 |
10以上 2 ・24 |
4 ・44 |
6 ・59 |
|
雌蛾の複眼を黒エナメルで塗りつぶしても、焼いても産卵に関する上記の明暗反応に変りがないが、複眼と共に腹部に黒エナメルを塗布すると、光線下においても暗の場合に近い蚕卵状態を示すから、腹部にも光覚があるらしい(426)。しかし、腹部だけに黒エナメルを塗っても、眼だけに塗った場合と同様に、正常な光反応を示す。
台紙の色は白黒何れでも産卵速度に違いがない(425,435)。
産卵の促進ではないが、割愛後、明るい場所において産卵を抑制しておき、これを暗所に移すと短時間内に比較的まとめて産卵させることができる(432,433)。
しかし、この方法は効果にかなりむらがあり、効果の明瞭でない場合や、初めから暗くして産卵を促進する方が却ってよい場合もある。これは、短時間の抑制ではその間に十分な数の成熟卵が集積しない場合や、反対に成熟卵が既に多数集積しているため、暗くしてその産卵を促進するだけで十分な場合などがあるためと思われる。効果のあった場合の1例を示すと(第82表)、割愛後直ちに暗くしたものでは産卵数歩合が60%に達するまでに4時間かかったが、初め3時間を明所においた後暗くしたものでは、暗くしてから2時間で産卵数歩合が70%に達した。
| 明暗調節 | 割愛後の時間別累加産卵数歩合(%) | 1蛾平均 産卵数(粒) |
||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7-24時間 | ||
| 全期間暗 最初1時間明 〃 2時間明 〃 3時間明 |
5 0 0 0 |
27 9 0 0 |
43 42 26 0 |
60 63 60 34 |
75 75 77 71 |
81 82 81 79 |
100 100 100 100 |
379 401 375 399 |
低温処理による産卵の促進とこの方法とを併用して、15℃、10℃、5℃、2.5℃、0℃、-2.5℃各60分間の低温処理を施した後、2時間明所におき、以後暗くして産卵させる試験も行なったが、対照(低温処理を施さずに、初めから暗くしたもの、および2時間明の後暗にしたもの)に比べて勝るものはなかったと云う。この原因には言及されていないが、低温処理は集積した成熟卵の産出を促進し、明保護は成熟卵の産卵を抑えてこれを集積させる作用を持つものと考えると、低温処理のあとで明保護をするよりも、明保護を先きにし、低温処理を後にする方が効果があるのではないかと考えられる。
c 産卵不良蛾
雌蛾は、割愛後、翌朝までの間に全産卵数の80-90%を産むのが普通であるが(第83表)、翌朝までに全然産卵しないものや極く少数しか産卵しないものがある。
| 蚕品種 | 試験場所 | 交尾歩合 (%) |
時間別産卵量歩合(%) | 総精選卵 量(g) |
||
| 第1日20時まで | 第1日20時から 第2日8時まで |
第2日8時から 第3日8時まで |
||||
| 日129号 | 新 庄 小淵沢 宮 崎 |
94 99 99 |
51 72 53 |
31 9 35 |
18 19 12 |
68 81 88 |
| 支129号 | 新 庄 小淵沢 宮 崎 |
96 100 100 |
74 81 75 |
11 8 16 |
15 11 9 |
73 106 88 |
大宮・滝沢(745)は日122号および支115号について、産卵数が造卵数の34%以下の少数産卵蛾の調査を行なったが、交尾、産卵中の保護温度を22℃、25℃、28℃に変化させてもその発現歩合に影響がなかった。少数産卵蛾の造卵数は正常産卵蛾に比べて少なくはないようである。顕著なのは受精卵歩合の低いことであった(第84表)。解剖してみると、交尾嚢や受精嚢中に精子の認められないもの、精子は認められるが交尾嚢の肥大していないものなどがあり、また正常精子束が少なく、無核精子を多く含んでいるものもあった。これらが、どのような受精卵歩合を示した個体を解剖してのことなのかは述べられていないが、少数産卵蛾には不受精卵が多く、その原因は雌蛾ばかりでなく雄蛾にもあることがわかる。
| 蚕品種 | 項目 | 調査した少数産卵蛾 | 正常産卵蛾 | ||||||||||
| No.1 | No.2 | No.3 | No.4 | No.5 | No.6 | No.7 | No.8 | No.9 | No.10 | No.11 | |||
| 日122号 | 造 卵 数 (粒) 産卵数歩合(%) 受精卵歩合(%) |
757 7 75 |
711 25 0 |
657 22 14 |
595 22 60 |
519 31 68 |
682 18 93 |
641 24 27 |
598 11 80 |
677 89 90 |
|||
| 支115号 | 造 卵 数 (粒) 産卵数歩合(%) 受精卵歩合(%) |
715 2 0 |
602 15 10 |
754 6 0 |
553 11 0 |
658 27 86 |
597 15 0 |
627 24 93 |
631 14 0 |
657 1 40 |
760 26 93 |
478 11 90 |
708 91 96 |
この少数産卵蛾の場合には、雌蛾にも相手の雄蛾にも生殖器の形態には特に異常がなく、卵管数の異常と少数産卵蛾の発現との間にも特に関係は認められなかった。しかし、卵管の中部に多数の卵細胞が塊状に集まっているものがあった。
不産卵蛾および少数産卵蛾は、一時冷蔵すると(50゚F、5時間)産卵するものが多いが、不受精卵が多く、再交尾させると不産卵蛾の約半数は受精卵を産み、少数産卵蛾は殆ど正常に近い産卵をすると云われている(264)ことは、これらが交尾の完全でなかった場合に多いことを示している。
三谷・渡会(545)は不産卵蛾の解剖調査を行ない、種々な生殖器異常を記載している。
d 産卵順序および産卵状態と蚕種の性状
1蛾の卵のうちで初めに産み出される卵は大きく、後のものほど小さいが、卵の大小と共にその性状に相違があり、晩産卵から孵化した次代蚕は飼育成績の劣る場合のあることについては既に述べた(Ⅱ2Ac)。これは実際問題に関係しているために、縷々試験が繰返えされているが、多くは、第1日目の産卵に比べて第2日目の産卵の飼育成績が劣ると云う結果で(629,693,895,968)、差がなかったと云う報告(738)は少ない。その影響の程度が種々な条件によって異なることは既に述べた通りである。
産卵速度のはやいものほど浸酸による孵化歩合が高い(381)と云われるのは、産卵が速いほど卵令の揃った卵が得られるためであろう。
受精率6.7%または7.6%と云うような高度不受精卵蛾区の受精卵でも、これだけを取出してみれば、孵化歩合にも飼育成績にも正常産卵蛾区の卵と違いがない(1000,1001)。
田中(1034)は、健康で若く性的に旺盛な雄蛾を交配すると雌卵を多く産み、また普通は雌卵と雄卵とが交互に産み出されるのであるが、雌卵が多くなるためにこの順位が乱れると云い、蚕の性染色体がZW型であることと関係があるように述べているが、蚕においては、交配する雄蛾によって卵の雌雄がきまるとは考えられない。
B 産み着け場所
a 産み着け面の角度
水平な面よりも、ある角度で傾斜した面に産み着けさせる方が産卵数が多いと云う結果と、これを追試したが水平産卵より勝ることはなかった(却って僅かながら少ない)と云う成績(706)とがあったが、藤本(98)は再びこの問題を取上げ、斜面に産卵させると、産卵の数は増加しないが、速度の早まることを報告した。
最も有効で個体変異の小さかった傾斜角度は、1蛾別産卵においては30°または40°であったが、40×30cm2の枠内に60蛾を入れた雑居産卵においては10°-40°が最もよく、その範囲内では差がなかった。斜面産卵の効果を、産卵数歩合が一定値に達するまでの時間の長短によって比較すると第85表の通りであるが、割愛後20時間内の産卵数および産卵数歩合を角度別に比較すると、第86表のように、産卵総数には殆ど違いがない。
| 産卵数歩合 (%) |
割愛後産卵数歩合が一定値に達するまでの時間(時間) | |
| 水平(0°) | 斜面(30°) | |
| 20 50 80 |
5-6 6-7 10-11 |
4-5 6-7 8-9 |
| 傾斜角度(°) | 産卵数(粒) | 体内残留卵数(粒) | 産卵数歩合(%) |
| 0 10 20 30 40 50 |
546 553 557 565 556 542 |
130 121 121 119 122 132 |
80 82 82 82 82 80 |
1蛾別産卵の成績によると、48時間の産卵では各区の産卵数歩合が更に接近する。傾斜角度が10°のときには上方へ移動する蛾が多く、50°では下方への移動が多く、30°では上下にほぼ同様な動きをするため、角度によって台紙面における餓の分布状態が異なると云う。
藤本は、斜面産卵によって産卵数歩合の高まるのは、蛾の産卵運動が促進されて産卵開始時刻が早まり、産卵速度もはやくなり、総べての蛾が揃ってよく産卵するためであろうと考えており、1蛾別と雑居とで、上記のように、幾分違いのあるのは個体相互の接触などの影響が加わるためであろうと云う。
藤本の結果において傾斜角度が40°以上になると効果の減少していることから考えると、最初に三谷が有効であるとしたのは約30°の斜面であり、岡村の追試は45°の斜面で行なわれているから、相反した成績の出たのは角度の相違によるものかも知れない。しかし、その効果が以前に考えられたような産卵数の増加ではなく、産卵の促進に過ぎないものとすると、午後6時から翌朝6時までの総産卵数をかぞえて結果を比較した岡村の方法では効果が検出できなかったとも考えられる。斜面産卵を採用しようとする場合には、この点をよく考えておく必要がある。
十万・片岡(299)も斜面産卵の効果を認めているが、最も有効な角度は20°で、30°がこれに次ぎ、40°では水平と大差がなく、40°では品種による差が大きかったと云い、藤本の結果と幾分違いがある。
大形の寒冷紗を懸垂してその面に産卵させる懸垂採種も以前に行なわれたことがあるが、岡村(706)の結果では、水平および斜面(45°)よりも産卵数が少なかった。しかし、その差は大きくない。
b 産み着け面の性質
| 産付け材料 | 1蛾当り産卵数 | 除外蛾 | |||||
| 調査蛾数 | 第1日目 | 第2日目 | 第3日目 | 合計 | 不受精卵 | 産卵数 300粒未満 |
|
普通台紙 糊引台紙 パラフィン紙 硫 酸 紙 寒 冷 紗 硝 子 糊引硝子 |
蛾 102 106 103 102 101 96 102 |
粒 561 531 524 535 563 478 467 |
粒 65 57 80 57 46 96 81 |
粒 5 9 15 5 6 5 5 |
粒 631 597 619 597 615 579 553 |
蛾 2 1 1 4 4 4 4 |
蛾 8 5 8 6 7 12 6 |
バラ種用の産み着け材料としては、多くの種類のものが試験されている。岡村(704)は、第87表の結果から、硝子および糊引硝子面には産卵数が少なく、除外蛾数の多いことに注目しながらも、試験区の差か否かは判定し難いとしている。しかし、牛込・服部(1128)の成績によっても、体内残留卵数(10時から15時まで交尾、翌朝8時まで産卵)の少ないものはセルロイド(10%)、晒木綿(10.8%)、寒冷紗膠付台紙*(11.5%)、渋紙(12.0%)、寒冷紗糊付台紙*(12.0%)、残留卵数の多いものが磨硝子(56.6%)、硝子(34.9%)であることからみて、蛾の脚が滑り気味の材料においては産卵がおくれ、産卵数も少ないことがわかる。 しかし、牛込らの成績において、硝子に次いで残留卵の多かったのがガーゼ(29.9%)、パラフィン塗布木板(24.9%)、フランネル(20.6%)であったことは、フランネルやガーゼのように毛羽立ったものも亦産み着け材料として不適当なことを示している。これらの材料においては、蛾の脚は滑らないが、毛羽のために動きにくいか、または卵を固着させにくいのであろう。(*註:台紙と云うのはおかしいが原著に従う)
水溶性の材料に産卵させておけば、卵の洗落しを機械化することができるのではないかと考えて、PVA(ポリビニールアルコール)フィルムを用いて試験を行なった際(873)の観察によると、15cm2ぐらいの大きさのフィルムを拡げただけで固定せず、その上で雑居産卵を行なわせた処、蛾の脚とフィルムとの付着の関係によるものと思われるが、蛾が体を廻しながら産卵するに連れてフィルムに渦巻き状の皺が生じ、その面に盛り上げたように累積卵を産んだ。産卵数も少なかった(未発表)。普通の場合の産卵を観察すると、尾端で1粒々々場所を探ぐって、前に産んだ卵の上には産み着けないようにするらしい行動を示すから、卵面と他の物体とを触知する覚感があるように思われるが、この結果からみると、次ぎ次ぎに産み出されてくる卵を固着させる尾端の動作と体の動きとのバランスがとれない場合にも累積卵を産むことが多いように考えられる。
また、このフィルムは蛾尿によっても溶けるために試験は失敗したのであるが、死ぬまでフィルム上で産卵を続けさせておくと、最後には尾端をフィルム面に粘着させたままになってしまう蛾が多かった。1粒産んでは尾端と体とを動かす動作が粘着のために制限される結果であろうと思われる。
これらの結果から、卵が固着し易く、蛾の動作の円滑に行なわれる産み着け面の望ましいことがわかる。バラ種用台紙に糊を塗り直して再製使用する場合があるが、産卵能率を上げるためには、これらの結果からみて、台紙の皺をよく伸ばし、蛾が円滑に動けるような面を作る必要がある。再製した台紙を使うと産卵が少ないと云うことは実際に耳にする処である。
c バラ種台紙用糊料
卵の洗落しを容易にするために台紙面に塗布する材料には、可溶性澱粉、生麩、ふのり、デキストリンその他種々なものが使用されてきたが(705)、最近ではCMC(Sodium carboxymethyl cellulose)糊が多く用いられる(638,639,874)。ポリビニールアルコールも使用することができるが、現在のところ、溶解性の点からCMCの方が使い易いように思われる(873)。しかし、これらの化学製品は従来の天然材料と違い、次ぎ次ぎと新らしいものが作られるから、これがよいときめることはむずかしい。また一口にCMCと云っても、種々な粘度のものがあって、使い易すさがそれぞれに違い、洗落しの難易も相違するから注意しなければならない。
天然糊料を使用していた当時は、黴の生えない糊が要望され、防黴剤を加えることも考えられたが、人工糊料を使うようになって、この問題は自然に解消した。しかし、これらの人工糊料は、それ白身は黴の栄養源にならないが、防黴力はないから、蛾尿や死蛾の体液などの付着している部分には黴が生える。
洗落したあとで卵同志の固着し合わない糊料の要望もあったが、卵同志の固着は大部分が卵自身の膠着物によるもので、台紙の糊が卵面に残っていて、それによって付着し合うことは殆どない。
累積(重積)卵は、台紙から洗落しても卵同志は容易に分離せず、相互固着卵として残ることが多い。しかし、あらかじめ累積卵を除去しておいても、洗落すと固着卵ができる。台紙と卵とは糊の層で隔てられているから、糊を水で溶かせば卵は離脱するが、卵殻面の膠着物は水で柔かくはなっても卵殻面に残っており、卵同志がこれによって付着し、乾かすと固まって、相互固着卵になるのである。
累積卵を除去して後に洗落した卵の相互固着卵50個をとって、どの面で付着し合っているかを調べたところ、
a 元来膠着物の付いていた面相互の付着 44
b 元来膠着物の付いていた面と付いていなかった面との付着 6
c 元来膠着物の付いていなかった面相互の付着 0
であった(873)。もし膠着物と関係なく付着し合うのであればbが最も多くなければならない。
膠着物による卵相互の固着を防ぐためにはホルマリンに浸漬して、付着し合うより前に膠着物を凝固させ、付着力をなくすることも行なわれているが、最もよいのは膠着物を除去することで、これには、洗落し後にクライト液に浸漬するのが最も簡単である(Ⅹ1C)。
CMC糊と従来の澱粉糊などとの洗落し上での著しい相違点は、澱粉糊の場合には、水に浸すと糊が膨潤し、卵は個々に台紙から離れ易くなり、洗落した後の紙面には糊の残っていることが多いのに対し、CMC糊の場合には、糊の層が膨潤すると、これが紙面からずるりと剥れたような形で卵を乗せて離れてくる傾向のあることである。糊の層が薄いと溶解するのでよくわからないが、糊を厚く塗って試験すると、膨れた糊層が寒天状になり、その面に胡麻をまいたように卵が付着して剥れてくるのがみられる。従って、糊の層が厚いほど卵はいたまないが、卵と糊との混合物ができて厄介である。これは従来の糊ではみられない特徴の一つである。
このような訳で、糊層の厚過ぎるのはよくないが、薄いと卵が糊層を通して紙面へ固着するので、落ちにくい。紙面への卵の固着は、洗落したあとの台紙を1%ぐらいのエオシン液に浸すと、固着の強かった部分には膠着物が残っていて、点々と赤く染まってみえるから、糊の厚さをきめる参考になる。
天然糊料は産地やマークが相違しても使用上の性質が全然違うようなことはないが、人工糊料の場合には、名前は同じCMCでも、粘度が違えば性質が著しく異なるから、購入に当っては、製造所、商品番号あるいは粘度などを正確に指定する必要がある。
Ⅴ 化性
以下の諸章で述べる蚕種の保護取扱いの主要な問題点は卵の越年性をどのように管理するかと云うことにかかっているから、その理解のために必要な化性の問題を説明しておく。
1 越年性の管理
蚕の卵には、産卵後一定の発生段階に達すると、外部の環境は発育に適当であるにも抱らず、胚の発育が一旦休止(休眠)して、ある程度の低温期間を経過しなければ発育機能を回復し難い性質のものがあり、このような卵は、自然状態においては、冬を越してから初めて発育を始めるので越年卵と呼ばれ、これに対して、休眠しない性質の卵は年を越すことなく孵化するので、不越年卵と呼ばれている。
総べての環境が一定であれば、蚕卵の越年性、不越年性は遺伝的性質によってきまる。春蚕の蛾が越年性卵を産む1年に1世代だけの種類を一化性、春蚕は不越年性卵を産み、その卵から孵化した蚕は越年性卵を産んで、1年に2世代を繰返えすものを二化性と云い、その他四化性、多化性などの種類がある。
厳密に云えば、越年する性質を持っている卵(産卵後の取扱いの如何によっては不越年性になる)を越年性卵、これが冬を越したものを越年卵として区別すべきであろうが、普通に越年種と云えば両方の意味に用いられ、そのときどきによって適当に解釈している。例えば、越年種の保護と云えば、越年性卵の産卵直後、未だ越年性の確定しないときからの保護を意味し、越年種の孵化と云えば冬を越した越年卵の孵化を指す。
越年性卵を黒種(くろだね)、不越年性卵を生種(なまだね)とも云うが、これを広義に用い、人工孵化法によって不越年にした蚕種を生種と呼び、人工孵化種にすることを“生にする”と云うような言葉の使い方も蚕種関係者の間では行なわれているが、関係者以外には通用しない場合が多い。
蛾になってから不越年性卵を産む(以下これを不越年卵性と呼ぶ)蚕は越年性卵を産む(越年卵性)蚕に比べて繭質が劣り、産卵量が少なく、不越年性卵は長く貯蔵することのできない不便がある。これに対して越年性卵は翌年まで貯蔵することができるばかりでなく、適当な時期に人工孵化処理を施すと不越年性に変えることができ、不越年に変えても繭質は劣らない。また催青法によって、二化性蚕の第1化目に越年性卵を産ませることができるので、日本では、現在、実用的には常に越年卵性になるように蚕を管理し、本来の不越年性卵を用いることがない。人工孵化種を生種と呼んでも蚕種関係者の間で間違いを起こさないのはこのためである。
産み出されたときの越年性卵は、越年性の方向は与えられているが、未だそれを完成していない状態にあるから(Ⅲ3Bc)、これを安全に越年させるためには越年性を完成させなければならない。これが越年蚕種の初期保護の眼目である。
越年性の完成は卵の休眠によって成立するのであるが、休眠した卵は、翌年の孵化させる時期に応じて休眠期間を調節し、予定の時期までに完全に休眠を解消させなければならない。これが越年蚕種の初冬期以後の保護法の主要題目である。
卵時浸酸は、越年性の確定するまでに卵に処理を施こし、休眠に入るのを中止させ、これを不越年性に変える方法である。不越年性に変わった卵は貯蔵に耐える期間が短かいので、産卵後40日以上もたってから孵化させたい場合には、休眠を幾分進めてから冷蔵すると、進めた休眠がある程度解消するまでに日数がかかるから、これによって貯蔵に必要な時を稼ぐことができる。これに浸酸を施こして孵化させるのが冷蔵浸酸法である。
このように、蚕種保護の重要問題はその多くが越年性の管理につながっている。
A 化性の変化
a 温湿度および光線による変化
温度による化性変化の方向は蚕の1世代を通じて同じではなく、3令を境にして逆転する(第88表)。
| 刺戟の時期 | 形質の変化方向 | ||
| 高 温 | 低 温 | ||
| 胚 子 期 | 4 眠 → 5 眠 不越年 → 越 年 |
4 眠 → 3 眠 越年 → 不越年 |
|
| 幼虫期 | 1-3令 | 4 眠 → 3 眠 不越年 → 越 年 |
4 眠 → 5 眠 越年 → 不越年 |
| 4-5令 | 越年 → 不越年 | 不越年 → 越 年 | |
| 蛹 期 | 越年 → 不越年 | 不越年 → 越 年 | |
| 蛾 期 | 越年 → 不越年 | 不越年 → 越 年 | |
しかし、その影響の最も強いのは催青中の温度で、二化性蚕種を25℃で催青すると、他の時期の温度の影響は殆ど覆われてしまい、蚕は越年卵性を獲得することができる(1139,1141)。
催青温度が化性に影響するのは胚に胸肢の発生する頃(Ⅵ2Be)から以後であるから、二化蚕の越年卵性を確実にするためには、少なくともこの時期から胚の頭部が着色し初める頃までを高温に保護する必要があるが、同じ日数だけ低温を作用させて不越年卵性蛾の発生歩合を比較すると、この期間のうちでも、後期になるほど低温によって不越年卵性蛾を出し易い(第89表)。
| 低温処理を 始めた時期 |
低温日数 (日) |
産卵結果 | ||
| 越年卵性蛾(蛾) | 不越年卵性蛾(蛾) | 不越年卵性蛾歩合(%) | ||
| A 胚に胸肢発生 | 1 7 10 15 |
294 240 185 0 |
0 2 14 215 |
0 1 7 100 |
| B 胚の体表に細毛発生 | 1 7 10 |
123 40 9 |
2 52 102 |
2 57 92 |
温度と共に化性に対する影響の大きいのは光線である(406,409)。木暮によれば、光線および高温が越年卵性蛾*を多くする方向に最も強く作用するのは催青中、これに次ぐのが稚蚕期(1-3令)で、3令にはその影響が小さくなり、壮蚕期(4-5令)には逆転して、不越年卵性蛾を多くする方向に働くようになる。蛹期においても壮蚕期と同じ方向に働く。有効な光の強さは温度によって異なり、20℃での催青中には0.0095F.C.、15℃催青においては0.0136F.C.が最低限界であるが、稚蚕期にはこれよりも更に高く0.077F.C.が最低限界であると云う。有効な波長の範囲は5,500Å以下(緑から青の方向へ)である。但し、この最低限界と云うのは、効果の認められるようになる最低限界の光の強さの意味で、越年卵性を確実にする光の強さの意味ではない。
*木暮は、温度が化性に影響するのは主として蛹および産卵後の時期で(Ⅴ2A)、催青中の温度および光は主として卵色に影響するものと考えているが、二化性種の着色卵は普通の場合には越年性卵と考えてよいから、ここでは着色性は越年性に等しいものとして記載した(83および152頁参照)。
越年卵性を確実にするためには、0.5F.C.以上の照度で、20℃以上の催青では毎日16時間以上、15℃催青では18時間以上の照明を、少なくとも胚の反転から気管の分化するまで(頭部着色の前)の間続ける必要がある。上記の時間照明すれば、1日中照明したのと同じ効果がある。
大造品種を24℃±1℃で飼育した場合に、その越年卵性を決定的にきめるのは幼虫期後期の短日処理(10時間明)であると云う報告もある。
催青中の湿度も亦化性に影響し(第90表)、低湿度ほど不越年卵性蛾を多くするが、催青温度の低い場合には、元来不越年卵性蛾歩合が高いので、湿度の影響が明瞭でない。
| 催青中の湿度 | 催青温度 | ||
| 17℃ | 21℃ | 24℃ | |
| 20-25% | 98% | 87% | 51% |
| 62-69% | 94 | 37 | 10 |
| 93-96% | 58 | 4 | 1 |
| 反転まで20-25% 以後93-96% |
89 | 24 | 2 |
| 反転まで93-96% 以後20-25% |
96 | 63 | 26 |
越年性管理の最も有力な手段に催青条件が挙げられるのは以上のような理由に基ずくのであって、越年卵性を確実にするための二化性蚕種の催青標準として温度25℃、湿度70-80%、1日16時間以上の明保護が採用されているのはこのためである。
b 遺伝性
化性は母親遺伝をする性質で、その優劣関係は一化性>四化性<二化性であると云われているが(1134,1136)、伴性遺伝も関係し(646)、単純なものではないらしい(616)。
同じ二化性と云われているものでも、品種によってその程度は著しく相違し、調査の時期によってもその表現にかなりの違いがある(第91表)。
| 系統 化性 |
1962年調査 | 1963年調査 | ||||||||
| 品種名 | 越年卵性蛾 歩合(%) |
不越年卵性 蛾歩合(%) |
混合卵蛾 歩合(%) |
調査蛾 数(蛾) |
品種名 | 越年卵性蛾 歩合(%) |
不越年卵性 蛾歩合(%) |
混合卵蛾 歩合(%) |
調査蛾 数(蛾) |
|
| 日本種 二化性 |
玉 光 神 光 瑞 光 日502号 日126号 日124号 |
5 99 100 93 66 95 |
84 1 0 0 0 0 |
11 0 0 7 34 5 |
19 79 113 30 38 38 |
郡 宝 し な の 昭 光 日122号 日124号 日126号 日127号 |
62 12 81 89 70 69 39 |
1 54 0 0 0 1 3 |
37 12 81 89 70 69 39 |
134 145 145 122 149 145 143 |
| 支那種 二化性 |
銀 白 栄 玉 支125号 支126号 支115号 支124号 |
41 75 0 6 1 6 |
48 21 100 44 71 51 |
11 4 0 50 28 43 |
106 147 91 123 163 124 |
春 光 た か ね 栄華(3眠) 栄華(4眠) 支115号 支124号 支126号 支127号 |
55 4 7 40 0 5 20 93 |
37 94 87 46 93 87 68 1 |
8 3 7 14 7 8 12 6 |
180 199 45 70 139 126 137 89 |
また、勝又(319)によれば、インドネシアの四化蚕に日本の二化蚕を交雑すると、第1代の卵が越年性になり、F2卵が、1蛾の中で、1/4は不越年I生、3/4が越年性に分離し、単因子遺伝を示したと云う。
化性は淘汰によっても変り易い。室賀(615)は二化性の日本種日新の1蛾区から出発して、越年卵性と不越年卵性との方向へ13代の淘汰を重ねた結果、20℃暗催青によって全部の蛾が着色越年性卵を産む系統と全部の蛾が不着色不越年性卵を産む系統とを分離することができたと云うが、梅谷(1112,1113)も、低温催青を行なえば完全に不越年卵性蛾ばかりを出していた二化性品種が、長年に亘って高温催青・人工孵化による系統維持を続けている間に、自然に一化性に近いものに変化していた例を報告している。
また、室賀(609)によれば、二化性種(正白および日新)の1蛾の卵を2分して、一半には高温明催青、他半には低温暗催青を施こし、孵化後同一環境の下で飼育して得た蛾を用い、高×低、低×高、高×高、低×低の交配を行なった処、高温母体のものは全部越年性卵、低温母体のものは全部不越年性卵を産んだ。これは期待通りの結果であるが、これらの卵の越年性のものには即時浸酸を施こし、不越年性のものはそのまま、共に20℃の暗催青を行なって次代を調べた処、高温母体(高×高も)の次代には不越年卵性蛾、低湿母体(低×低も)の次代には越年卵性蛾が多かったと云う。この場合は、同一母体の卵から出発しているので、遺伝的な分離の結果ではなく、生理的な現象と考えなければならない。
c その他の原因による化性変化
長谷川(153,157)は化蛹後2、3日の蛹に硝酸ウラニウム10M水溶液を0.05ccぐらい注射すると、越年卵性のものが多数の不越年性卵を産むようになることを発見した。化蛹2日目に注射した場合の卵を産卵順序によって初、中、晩に分けると、初期の産卵は総べて不越年性卵、中期のものは不越年性卵と越年性卵との混合、晩期の産卵には越年性卵が多く不越年性卵が極めて少なかった。これは、ウラニウムが、注射当時に未だ越年性に方向付けられていない卵のうちのある程度発育しているものにだけ作用するためであって、化蛹5日目に注射すると殆ど全部の蛾が越年性卵を産み、混産蛾においては、初期の産卵が全部越年性であった。水および体液に難溶性の燐酸ウラニウムを化蛹初期の蛹に挿入しても効果がなかったから、上記の結果は放射線の影響によるものではないと云う。
食道下神経節を摘出あるいは移植すると化性の変化することは後に述べる。福田(109)は、不越年卵性蛹の気門のいくつかをエナメルで閉ざすと越年性卵を産んだと報告している。エナメルに少量のガソリンを加えると効果があるらしい。福田(116)はまた越年卵性蛹の皮膚に傷をつけたり、ある種のホルモン剤、卵黄、空気、ガラス片、繭片などを注射または挿入したりすると不越年性卵を産み、不越年卵性蛹に同様な処置をすると越年性卵を産むことが縷々あると云い、これらの処置が食道下神経節の分泌活動を変化させるのであろうとして、ストレスによる化性変化と呼んでいる。
吉武(1205)は、越年卵性蛹に硝酸ウラニウム、KCl、種々な酵素阻害剤などを注射すると不越年性卵を産むが、不越年卵性蛹にKCl、クエン酸ナトリウム、アジ化ナトリウムなどを注射すると越年性卵を産むと報告している。
西郷(788)は、二化性蚕種の催青中に炭酸ガスを接触させると不越年卵性蛾が多くなることを報告した。催青温度19℃-20℃のときに、炭酸ガス濃度3-5%で処理するのが最も有効で、付属肢形成期から孵化期までの間において接触期間の長いほど効果がある。例えば、19℃(支106号は20℃)、炭酸ガス濃度4%、催青期間中接触の場合の不越年性卵蛾歩合は日110号24%(無接触3%)、乞食96%(同64%)、支106号55%(同19%)であった。
不越年卵性の強い大造種の蚕を一化性または二化性系統の蚕と混合飼育すると大造の越年卵性が高まると云う報告がある(443,444)。原因としては、一化性蚕の残桑を大造に与えると越年性の強まる傾向があるとか、混合育によって大造の食桑が活発になるためではないか、などと考えられているが、確実なことはわかっていない。
これらの結果は、何れも最終的には食道下神経節の分泌活動に帰せられるものではあろうが、種々な誘因がどのような機構でそれにつながるのかは未だ明かにされていない。ただ極めて多くの条件が化性変化の誘因になり得ることは明かであるから、蚕の一生を通じて、異常な環境や取扱いを避ける注意が大切である。
B 休眠
a 休眠の完成
蚕卵の越年性の完成は卵の休眠によって成立するから、休眠の調節は越年性管理の重要な部分を占める。催青、飼育、上蔟、発蛾の全期間を通じて、保護条件を越年卵性を高める方向に調整しても、実際に越年性の確定するのは産卵後である。
越年性への方向付けの不十分な卵を越年化させるには産卵後暫くの間低温(15℃)に保護するのがよいが(Ⅲ3Bc)、15℃に保護しただけでは越年性の方向はきまっても、完成はしない。
蚕の越年性卵は、休眠に入ると、ある期間低温に接触しなければ正常な孵化機能を得ることができないが、高温においたままでも全然孵化し得ない訳ではない。産卵直後から15℃以上の種々な温度に保護しておき、その温度での越年性卵の孵化をみると(第92表)、15℃での孵化が最も早い。これは休眠が完成しないまま越年性を失なうためである。 同様に20℃においても25℃に比べて休眠が不安定である。30℃においては全然孵化せず死滅している。
| 保護温度 (℃) |
日本一化 | 支那二化 | 日本一化×支那二化 | 支那二化×日本一化 | ||||
| 孵化歩合 (%) |
孵化の日数 (日) |
孵化歩合 (%) |
孵化の日数 (日) |
孵化歩合 (%) |
孵化の日数 (日) |
孵化歩合 (%) |
孵化の日数 (日) |
|
| 30 25 20 15 |
0 6 57 93 |
- 245-393 139-439 127-204 |
0 11 68 93 |
- 250-373 138-396 121-170 |
0 32 79 94 |
- 245-391 138-456 121-192 |
0 36 83 99 |
- 255-378 159-456 121-197 |
この表は、休眠のほかに生理障害の加わった孵化成績を示していると考えられるから、例えば、30℃においても、その期間を短かくすれば、生理障害を与えずに休眠を完成させ得るとは思われるが、第51表をみると、28℃に3日間おいただけのものでさえ、25℃においたものに比べて遙かに劣る結果が出ている。従って30℃は蚕種にとっては正常をはずれた温度と考えられ、休眠の完成には25℃を中心とした温度が実用上最も安全なことがわかる。
産卵後、越年性卵を25℃で引続き保護すると、90日ぐらいまでは、25℃期間の長いほど孵化機能を得るのに要する低温日数が長くなるが(第51表)、これが、休眠の完成に90日かかることを示すものとは考えられない。
休眠に伴なう蚕卵の生理的変化をみると、25℃内外の温度においては、産卵後、
核分裂の認められなくなるのが 約4日(386)
卵汁のpHの最低値になるのが 約10日(965)
卵黄細胞の移動の完了するのが 約10日(928)
チロシナーゼ作用の最低になるのが 約14日(687)
グリコーゲン含量の最低になるのが 約30日(81)
炭取ガス排出量の最低になるのが 約30日(872)
であるから、生理作用の最低になる時期を目標にするならば、休眠は産卵後30日以内に完成するものと考えられる。
また、蚕種保護の面からみても、25℃の期間が30日を超えると越年種の孵化歩合が低下するから(第93表)、休眠完成のための高温保護期間は30日以内にとどめるのがよい。但し、これは原種の場合で、交雑種においては25℃期間が50-60日になっても障害のない場合が多い。
| 25℃日数(日) | 供試卵数(粒) | 孵化粒数(粒) | 孵化歩合(%) |
| 34 54 85 117 146 162 25℃に放置 |
4,189 4,314 4,179 4,466 4,685 4,606 4,419 |
3,696 3,478 3,234 1,286 1,013 596 6 |
88 80 77 28 21 12 0 |
大野(766)は、25℃保護90日ぐらいの頃が休眠解消(活性化)に要する低温保護日数の最も長いときで、高温日数が100日を超えると却って活性化し易くなると報告したが、これは第51表および第94表においても明かである。大野ら(764,1118,1119)はまた、30-60日、特に35-40日頃に、その前後に比べて冷蔵または洗落しの機械的刺戟によって幾分活性化し易い時期があると云う。材料に用いた蚕品種は異なるが、河合(349-351)の洗落し試験においては、50-60日で孵化の多くなることは認められるが30-40日にこのような時期のあることは認められない。しかし、このような考えで高梨らの成績(第51表)をみると、省略した高温20日、50日、70日の成績をも加えた中で、高温30日で冷蔵したものの孵化が冷蔵日数の短かい時期において幾分勝っている。偶然の一致であろうとは思われるが、もしこのようなことが確認されれば休眠の生理上問題である。
b 休眠の解消(活性化)
休眠卵の活性化のために最も有効な温度は5o- 7.5℃であるが、蚕品種によ
って幾分違いがあり、活性化が、5℃よりも幾分高めの温度において速く5℃以下ではおくれるものと、5℃以下でもかなり速いものとがある(685,936)。従って、活性化の早さだけからみれば、品種によって、5℃と7.5℃との何れがよいとも云われないが、蚕種保護において一定の低温を用いて休眠卵を活性化させるのは、活性化を促進すると共に、早く活性化した卵の発育をできる限り抑え、その間におくれているものの活性化を進め、全体の発育を揃えるのが目的であるから、活性化の目的にそう限り温度の低い方が望ましく、この点からは5℃が7.5℃に勝る(第102表)。
2.5℃は活性化の促進においては5℃に劣るが、卵の生理作用を抑え、活力の低下を防ぐ点では勝っているので、蚕種の貯蔵を兼ねて活性化の不足を補なう温度として用いられる(第94表)。
| 低温の種類 (℃) |
孵化歩合が80% 以上を示す冷蔵日数 |
冷蔵前の25℃日数(日) | |||||||||
| 1 | 5 | 10 | 20 | 30 | 60 | 90 | 120 | 150 | 180 | ||
| 7.5 | 最 短 日 数 最 長 日 数 期 間 |
30 120 90 |
60 150 90 |
60 150 90 |
60 150 90 |
60 150 90 |
60 150 90 |
90 150 60 |
60 150 90 |
60 120 60 |
60 120 60 |
| 5.0 | 最 短 日 数 最 長 日 数 期 間 |
30 180 150 |
60 210 150 |
60 210 150 |
60 210 150 |
60 210 150 |
90 210 120 |
90 210 120 |
60 180 120 |
60 120 60 |
60 120 60 |
| 2.5 | 最 短 日 数 最 長 日 数 期 間 |
60 180 120 |
90 180 90 |
90 210 120 |
90 180 90 |
90 210 120 |
120 240 120 |
150 210 60 |
120 210 90 |
90 180 90 |
90 150 60 |
以上は産卵後25℃で1日以上経過して、卵に越年性卵の特徴の現われ初めた(Ⅵ2Ba)頃から後の卵の活性化について述べたのであるが、河野(450)によれば、これより若い、産卵直後から1日目頃までの間の越年性卵を種々な温度で一定期間保護した後、25℃で催青すると、活性化が最も速いようにみえ、初期の孵化合が高いのは15℃保護の卵である(第95表)。
| 処理温度 (℃) |
処理後の期間別孵化歩合(保護温度25℃) | |||
| 55日間(%) | 56-220日間(%) | 220日以後(%) | 計(%) | |
| 5 7.5 10 15 20 25 |
0 0 0 15 0 0 |
0 0 3 6 3 0 |
32 40 31 11 42 51 |
33 40 35 33 45 52 |
種々な実用品種の越年性卵を産卵直後からこれらの温度に保護し、その温度における孵化を調べても、矢張り15℃における孵化歩合が最も高く、20℃、25℃
の順でこれに続き、10℃においては太平が0.1%孵化したに過ぎず、5℃および7.5℃においては孵化したものがなかった。卵を解剖してみると(大造)、0℃および5℃においては何時までたっても胚の形成が認められず、10℃においては6ヵ月後に漸く幼虫体が完成したが、15℃においては4ヵ月で孵化がみられた。河野はこの結果から、産卵後1日以内の若い卵の活性化促進に最も有効な温度は15℃であると考えているが、低温においては胚の形成が悪いこと、第95表において孵化歩合合計の最も高いのが15℃ではなく25℃で、次ぎが20℃であることなどを考え合わせると、低温と高温との中間の15℃においては、活性化と胚形成との条件が両立する結果として孵化が早いのではないかと考えられる。
一定の温度に保護してある卵については、品種による相違などはあっても、第51表や第94表によって、現在の活性化程度が大約察知できるが、実際の蚕種保護においては、種々な自然温度に遭遇するので、活性化程度の察知がむずかしい。室賀(621)は、この点につき、各種の温度の活性化効果を同一基準で評価することができれば蚕種取扱い上便利であると考えて、抑制質消耗係数を発表し
た(第96表)。
| 温度(℃) | -2.5 | 0 | 2.5 | 5 | 7.5 | 10 | 12.5 | 15 | 17.5 | 20 |
| 抑制質の 消耗係数 |
0.00 | 0.40 | 0.77 | 1.00 | 1.11 | 0.77 | 0.61 | 0.41 | 0.17 | 0.00 |
これは、蚕種の活性化は母体から卵に伝えられた抑制質が消耗する結果として起こるものであると考えて名付けたのであるが、具体的に云えば、5℃に1日おいた場合の活性化の進み方(抑制質の消耗)を1とすれば、15℃の1日では0.41しか進まないから、5℃1日と同じ活性化程度にするためには、15℃には1/0.41=2.4日強おく必要があり、また15℃に10日おいた卵は5℃に4.1日おいたのと同じ程度に活性化が進んでいると云うのである。
このような方法で大凡の見当をつけることは便利であるが、活性化の進み方は温度ばかりではなく、洗落しの刺戟などによっても影響を受け、また品種による反応の相違もあるから、これだけに頼ることはできない。
蚕の卵は休眠状態で冬を越すと考えられていることがある。例えば、梅谷(1106)は昆虫の卵態越冬を5群に分類し、蚕はアレイ形の胚の状態で越冬する第2群に属し、産卵後7-10日の前休眠期間を経て休眠(自己休止)に入り、その後2月頃までが休眠期であるとしているが、地域的な相違もあるが、2月以前に孵化可能な卵の現われることは周知の通りである。蚕種に委しい梅谷博士がこれを御存知ない筈はなく、実用上の催青期間内に全部の蚕種が孵化し得るまでに至っていないことを表現されたに過ぎないが、文字通りにこれを解釈すると間達いのもとになる。もし蚕種が完全な自己休止の状態で冬を越すのであれば取扱いは比較的簡単であるが、完全休眠卵に混じって、完全に孵化し得る卵、孵化にまでは達し得ないがある程度まで胚の発育し得る卵などが初冬期頃から出初めるために、早く活性化したものの活力を低下させず、おくれたものを追付かせて、発育を揃えることが蚕種の冬期保護上最大の課題になっている。
西郷(788)は、休眼中の卵に炭酸ガスを接触させると活性化を早めると報告した。
2 休眠の生理
A 抑制質
蚕卵の休眠現象を説明するために種々な仮説が提出されているが、その中で、抑制質の仮定(Ⅲ3Bc)は蚕種保護の技術と結び付いて蚕糸関係者の常識になっており、最初、渡辺が便宜上仮定したものが実在のものとして受取られている場合も極めて多い。
木暮(406,409)は、着色卵のうちの不越年卵(再出卵)および不着色卵中の越年性卵の割合が何れも催青中の温度および光線の影響によって変化しないこと、二化性蚕および一化性蚕においては、蛹期の保護温度が30℃の場合に20℃の場合よりも越年性卵の多いこと、四化性蚕においてはこの関係が反対であること、および産卵後2日間の温度が越年性の決定に強く影響することなどから、第1越年性物質(着色性物質。渡辺の抑制質と同じと考えられる)および第2越年性物質(越年性物質)の存在を仮定した。第1越年性物質は越年性にも関係するが主として卵の着色に関係するもので、高温・明催青で生産される。最も多く作られるのは反転から気管形成までの血球分化の時期で、以後3令の終りまで次第に生産が減り、4-5令および蛹の時期においては高温・明の条件下で破壊されるようになり、この時期の低温によって生産の促進されることもない。第2越年性物質は卵の着色にも関係するが主として越年性の決定に関係する。蛹期の高温によって作られ、産卵後の高温によって破壊される。一化性蚕には第1、第2ともに多く、二化性蚕には第2は多いが第1が少なく、四化性蚕には両者ともに少ないと考えている。
梅谷は化性決定素を考えたが、これは母体から卵に伝えられる酵素である(1101)。不越性卵はこの酵素の活性によって卵細胞質の生化学的活動が持続している卵であり、この酵素が一時的に不活性になって、卵細胞質の生化学的活動の停止している卵が越年卵であると考えている。この仮定は実用との結び付きは抑制質の仮定のように一般的ではないが、この考えから出発した硬性卵、軟性卵と云う言葉は越年性の保護に関して縷々使われている。硬性卵は、産卵後、高温に十分な日数接触して卵細胞質の生化学的活動が停止し、栄養質が不溶解性になっており、低温処理による生化学的活動の回復が緩慢で、栄参質の可溶性になるのがおそい卵と云われ、軟性卵は、高温接触日数が少ないため、低温処理リこよって卵細胞質の生化学的活動が急激に回復し、栄養質の可溶性になるのが速い卵であると説明されている。生化学的活動状態とは云っても具体的に物質の動きを証明している訳ではなく、水分量の変化と結び付けて解釈しようと試みているが(Ⅵ3A)、別の言葉で云えば、硬性卵とは休眠の完成した卵、軟性卵とは休眠完成の不十分な卵である。
抑制質の仮定は蚕種保護技術の確立上、一般に理解し易い説明の根拠となり、実際上に貢献したことは極めて大きいが、越年性卵中に胚の発育を抑える作用を待った物質の存在を証明した報告はなく、これと休眠ホルモンとの関係についても見解は一致していない(Ⅴ2C)。
胚培養によってこの点を調べてみると、(A)抑制質のない筈の卵から胚だけを取出し、同様な卵を磨砕して生理食塩水で抽出した培地で培養すると胚が発育する(930,931,949-951)。(B)産卵後2、3日たった休眠性卵の胚を同じ時期、および、(C)抑制質のない筈の卵で作った培地で培養しても、または(D)抑制質のない筈の卵を産卵後2、3日の体眠性卵の抽出液で培養しても、同様に発育するが、(E)産卵後30日以上もたった完全休眠卵の胚は同じ時期の卵の抽出液は勿論、抑制質のない筈の培地においても育たず、(F)完全休眠卵抽出液培地においては、抑制質を持たない筈の卵の胚も発育しないかまたは発育が非常に悪い。
(A)の発育するのは当然であり、(B)および(D)が同じように発育するのは、培地を作る操作の間に抑制質が破壊されたためであると考えられないことはない。母体から卵に伝えられた抑制質は初めは原形質中に存在し、このときは外部からの刺戟によって破壊され易く、従って人工孵化も可能であるが、やがて抑制質は胚に移行して胚を完全に休眠させ、胚に移った抑制質は、も早、外部からの卵を殺さない程度の刺戟では破壊されなくなると云う考え方(Ⅷ3A)もあるが、抑制質が胚の中に移った筈の完全休眠卵培地において、(F)のように(B)、(D)よりも発育が悪いことは、仮りに抑制質の1部が原形質内に残留しているものと考えても説明ができない。
(F)の場合に発育が悪いのは培地の中に抑制質が残っているためかも知れないと考えて、(A)や(B)についての経験から十分に発育可能と思われる範囲内で培地を稀釈しても発育は改善されず、ラクトアルブミンの加水分解物やアミノ酸を添加すると胚の発育がよくなった。これらの結果は、(F)の発育が悪いのは抑制質の存在よりも、利用し得る状態で培地中に存在する栄養物質の量または質の問題であることを示している。
休眠卵が活性化するのは、胚がまず休眠から醒めて生化学的な活動を始めるのか、それとも、まず卵黄質の状態に変化が生じ、これが胚の発育を誘導するのかと云う問題をめぐって、激しい論争の行なわれたことがある(549,550,1101,1102)。
上記の卵外培養の成績によれば、休眠卵においては胚も卵黄も共に発育に不適当な状態に変っていることがわかる。
梅谷(1104,1123)は、休眠卵に非休眠卵の卵黄を移注すると休眠卵の胚が発育し、トリプシンを注入しても発育の開始されることをみて、蚕卵が休眠から醒める際に、まず非可溶性蚤白質が可溶性に変化する証拠であると考えたが、食塩水を注入したり、卵を針で刺すだけでも休眠胚の発育の始まることがわかり、これは物理的刺戟によって卵細胞質の生化学的作用が活性化するのであると説明した。なお、休眠卵の卵細胞質を不越年性卵に移住しても、これを休眠させることはできなかった。
卵黄は単に栄養物質が卵内に充満しているのではなく、卵黄分割(Ⅵ2Ba)の後には、生活機能のある卵黄細胞に含まれて存在するのであるから、これを溶解する梅谷の生化学的作用も卵黄細胞の機能を通じて働く筈である。
卵から卵殻だけを除去し、生きた卵黄細胞群および漿膜に包まれた胚をそのまま適当な培地で培養すると、非休眠胚に比べれば劣るが、完全休眠胚も発育して剛毛を生じ、頭部の着色するまでになることが珍しくない(941)。これによってみると、上に述べた胚だけを取出した場合に完全休眠胚の発育しなかったのは、培養技術が未熟なために、発育開始のおそい完全休眠胚を十分に生存させ得なかったのであるうとは思われるが、また卵黄細胞をすり潰して、正常な生活機能を破壊した培地の中では、活性化その他の生理作用が進行しにくいのではないかとも考えられる。しかし、現在この方面から梅谷の考えを支持する証拠は得られず、バッタなどにおいては、卵黄から完全に分離した胚の組織片が生理食塩水中でも活性化すると云われている。
B 休眠に伴なう卵内の変化
前節で述べた培養実験の結果から、休眠卵においては、胚や卵黄の生理機能がただそのまま停止しているのではなく、非休眠のものとは著しく異なった状態に変化していることが考えられるが、このことは休眠卵における糖代謝の研究によって実証された。
休眠に伴なう蚕卵内のグリコーゲンの変化については、古くから多くの研究があるが(273-276,468,536-538,548,549,672,1169)、これらの多くはただその増減を記載するに止まり、休眠初期にみられる著しい減少を生活のために消費されるものと考え、初冬期の増加は蛋白、脂肪などから新たに作られるものと考えるのが普通であった。
茅野(76-79,81-85)は、蚕卵が休眠に入る際には、これまで昆虫においては全然考えられなかった代謝径路によってグリーコーゲンがグリセロールとソルビトールとに変化し、休眠が醒め初めるとこれが再びグリコーゲンに戻ることを明かにした。即ち、休眠は単なる生活機能の停止ではないのである。
休眠に伴なう蚕卵の生理作用の変化については、その他、酵素(40,78,84,187,191,399,664,686-688,818)、呼吸(683,684,781,785,872,893)、水素イオン濃度(148,593,594,625,965)、電気泳動像(666)、卵汁の屈折率(213)、水分(467,671)、3・ヒドロキシキスレニン(244,354)、燐化合物(80)、脂肪(670)などに関する多くの報告がある。しかし、これらの変化が休眠の完成または解消の本質に関係した動きなのか、平行現象であるのか、または休眠の結果であるのかは明かでない。
この点に関して興味のあるのは室賀、西郷らの報告である。室賀(608)は、日新種の産卵直後の不越年性卵を炭酸ガスを含む容器中に2、3日間密閉すると漿膜の着色する卵が多数に生じ、炭酸ガス濃度が2.5%に達するまでは濃度の高いほど着色卵歩合の高い結果を得た。この着色は赤褐色で、不越年性卵を11℃に20日間、または13℃に15日間保護した場合に生ずる越年性卵に似ていた。産卵したのは6-8月であったが、これを自然温度で保護すると越冬前に不時発生をして催青死卵になるものが多く、翌春になって孵化したものは1頭もなかった。人工越冬を施すと最高の9.39%の孵化を示した。即ち、極端な軟性卵と考えられるような卵が得られた訳である。
西郷(780,784)はこれを追試し、炭酸ガス濃度、処理時間および処理時期と着色との関係を調べ、最も着色を起こし易いのは、産卵後25℃で1-2日目の、胚の頭尾が、窒素、酸素によっても着色することを認めた。炭酸ガス処理によって生じた着色卵は20℃に50日間おいても孵化しなかったが、酸素消費量、呼吸商および青酸による呼吸阻害などは何れも正常越年性卵の産卵後25℃、2日目に相当する状態に止まったままで、正常休眠卵のような青酸に阻害されない呼吸に移行することがなかった。呼吸商は、正常の越年性卵においては0.70から次第に大きくなり、産卵後2、3日で、0.74、
4日目に0.76になったのに対し、炭酸ガス接触による着色卵においは0.68から増大して4日目に0.72となり、7日目以後は0.74に一定した(Ⅵ3Bb)。しかし、卵黄細胞の移動(Ⅵ2Ba)は行なわれる。
これらは呼吸障害の結果だけによるものではなく、炭酸ガスの何らかの作用によることも大きいのではないかと思われるが、産卵直後の不越年性卵を暗黒下でCO288:O212 の混合気体中に入れておくと、漿膜が赤褐色に着色し、卵黄細胞の移動、休眠期の形態での胚の発育停止が起こり、チトクロームオキシダーゼの活性およびグリコーゲン量が低下し、5、6日で休眠卵における値とほぼ等しくなった。これに対し、同じ処理を同所で行なうとこのような変化が起こらなかったから、上記の結果はチトクロームオキシターゼ作用の阻害の結果と考えられる(786)。ただ、14日間で混合気体中から出すと、直ちに発育を始める卵もあれば、休眠状態を続けるものもあって一定しないから、この酵素系の阻害は休眠を誘導する条件ではあるが、十分な条件ではないのであろうと云う。
西郷の報告は簡単な講演要旨に過ぎないが、非休眠卵の呼吸においてはチトクロームオキシダーゼが主な末端酵素として働いていると考えられること(684)、および、茅野(83)が、蚕卵内にはグリセロールおよびソルビトールの生成に関与する四つの脱水素反応があり、このための脱水素酵素は非体眠卵中にも存在するが、休眠卵においてだけ上記の多価アルコールが作られるのは、休眠の初めにある物理化学的変化が起こって末端酵素系における電子伝達をブロックし、その結果NADH2およびNADPH2が蓄積して、反応をこれらの多価アルコール形成の方向に進めるのであろうと考えていることなどと関連して興味がある。
不越年性卵を越年化させる最も普通の方法は産卵直後からの低温保護である。木暮(409)によれば、13℃に15日間保護すると最も効果があり、低温期間がこれより良くても短かくても効果が減少し、産卵後2日までは未だいくらか効果が認められたが、3日以後の低温保護(15日間)は無効であった。室賀(616)は、低温保護によって不越年性卵が越年化するのは、胚の発育がおくれるため、卵を濃く着色し得ない程度の抑制質でもその発育を抑止して、越年化させることができるのであろうと考えている。
再出卵防止のための低温保護(Ⅲ3Bc)もこれらと同じ意味のものである。河合(355)は、産卵後1週間27℃に保護した蚕卵と15℃においたものとを比較して、却って前者に再出卵の少なかった場合を報告し、この場合の材料が多化性に近い大造系統であったため、普通の二化性種とは温度に対する感受性が異なり、27℃を越年性を安定させる温度として感じたのであろうと考えている。
電子顕微鏡で観察すると、休眠に伴なって胚細胞内のミトコンドリアの形態が変化して、著しく長形になることが注目されているが(554,702)、岡田(702)は休眠卵においては、小胞体が顕著な同心円状に配列すること、およびリボソームの減少することをも観察している。また、培養した卵のH3-チミジンおよびH3-ウリジンの取込みを調べ、DNA合或は休眠に入ると直ちに停止し、低温で休眠を解消させても温度を上げなければ合成の再開しないこと、RNA合成および蛋白合成は休眠に入っても長い間停止せず、100日ぐらいで非常に少なくなるが、140日では回復し始めることをみている。
RNA合成が回復しても胚は直ちに孵化機能を得る訳ではなく、休眠は徐々に解消する。
RNAの取り込みの再開は胚の後端部から始まり、福田らの観察した休眠解消の不完全な胚が特異な発育をする現象(Ⅷ4)と関連があるように考えられると云う。これは越年性卵における核分裂の消長(386,945)や胚培養の結果(Ⅴ2A)とも考え合わせて興味のある結果である。
C 休眠ホルモン(休眠要因)
越年卵性の蚕においては、食道下神経節から越年性を決定する物質(長谷川の休眠ホルモン、福田の休眠要因)が分泌され、卵巣内で発有中の卵に作用してこれを越年化させることが福田(111,112,114,117,132-136)および長谷川(158-162,164,166,167)によって初めて明かにされた。これは我国で殆ど同時に発表された世界的研究で、これによって蚕の化性に関する考え方が一変した。ただ、この要因の分泌が脳の支配を受けており、多化蚕が不越年性卵を産むのは、神経節から要因が体内に出るのを脳が抑止しているためであると云う考え(福田)と、食道下神経節のホルモン分泌は脳の支配を受けないと云う考え(長谷川)とはなお一致していない。
食道下神経節の休眠ホルモンは神経分泌細胞から分泌されるものと考えられるが、小林(400,401)によれば、神経分泌細胞と判断される細胞は16μぐらいの小形のものから60μに近い大形のものまで種々あって、1個の食道下神経節につき80-100個ぐらいかぞえられ、品種によってもこの数に相違がある。同一母蛾の産んだ卵を2分し、一半は越年性卵を産むように、他半は不越年性卵を産むように処理し、その5令幼虫について神経分泌細胞を比較した処、数には殆ど差がなかったが、長径30μ以上の大形のものの割合は、不越年化処理を施したものよりも越年化処理を施したものに多く、2倍に近かったと云う。
これに対し、最近、福田・竹内(138,139)は、休眠要囚は、蛹および蛾の食道下神経節において、前端から約2/3の部分で腹面に近く、中心線を挟んで相対する1対の大形細胞(DF細胞)だけから分泌されると云う実験ならびに観察結果を報告した。
休眠ホルモンの抽出、精製は長谷川によって進められているが(163,164,168)、未だ精製品は得られていない。しかし、精製過程の抽出物を注射してホルモン作用に関する多くの実験が行なわれ、蚕種の立場からも興味のある結果が得られている。不越年卵性蛹について試験した結果によれば、化蛹後(27-28℃)3日目頃の注射が越年性の誘導に最も有効で、注射後5日間15℃に保護しておくと効果が一層大きかった(169)。これは、化蛹後、種々な時期の休眠性蛹から食道下神節節を除去し、蛾になってから産む越年性卵歩合を調べると、化蛹後3日までに除去したものは休眠性卵を全然産まなかったことと時期的に一致している(1173)。
越年性卵の漿膜細胞の着色は、主としてトリプトフアン系色素で、これにチロシン系色素が関与しているのであろうと云われ(247,371,373)、メラニン系色素は存在しないらしい(717)。不越年性卵の漿膜が着色しないのは、母体内で生産されたキヌレニンまたは3・ヒドロキシキヌレニンの卵内への透過が妨げられるのか(246,315,354-357,371,636,1203-1206)、あるいは透過しても吸着されないのではないか(1207)と考えられているが、食道下神経節の摘出および休眠ホルモンの注射実験によって、休眠ホルモンが卵巣内の3・ヒドロキシキスレニン量に影響していることがわかった(1170,1174)。糖および脂質の代謝も休眠ホルモンによって規定されており、越年性卵にグリコーゲンの多いのもこのためである(171,1171-1174)。
休眠ホルモンは5令壮蚕、蛹および蛾の食道下神経節のほか、蛾の頭部からも抽出され、雌よりも雄に争い。卵からは未だ抽出されていない(164,169,170)。福田・河野(137)は卵内にも休眠要因またはこれに関係のある物質が存在するものと考えている(Ⅷ4)。
休眠ホルモンが卵巣(特に包卵細胞)の遺伝子に作用してm-RNAの合成を促がし、これが出発点となって越年化への種々な生理作用が次ぎ次ぎに進行する(172)のであれば、このホルモンのほかに特別に胚の発育を抑制する物質の存在は必要がないように考えられるが、卵の着色と越年性とが平行するのは、3・ヒドロキシキヌレニンおよび抑制質の卵内への移行が共に化性ホルモンによって規正されるためであるとして、ホルモンとは別に抑制質の存在を考える立場もある(356,1204)。
食道下腺を化性に関する内分泌器官であるとする報告もあったが(551)、これは福田(109)および長谷川(160)によって否定された。
1 受精前の蚕卵の形態
家蚕の卵は、普通、偏平な小楕円体で、大きさは大約、長さ1.3mm、巾1.0mm、厚さ0.6mm(第42表)であるが、品種(697,982)、栄養(Ⅱ2Ac)、蛹の保護温度(Ⅲ3Bb)その他種々な条件によって変異をあらわす。また遺伝的な畸形も知られている(75,1037)。
産卵当時は乳白乃至黄色である。これは主として卵殻および卵黄膜を通してみえる卵黄の色によるものであるが、卵殻が着色しているものではその色も加わっている。
卵殻は最外層の卵膜で、均一な構造ではなく、多くの層を識別している報告もあるが、普通は外層、中層、内層および最内層に分けている(Ⅰ2Abii)。
卵殻の厚さは品種によって相違するが(568,697,916)、卵形を大きくする条件の下では卵殻も厚くなる。また卵の部分によってもかなり厚さが異なる(697)。最も厚いのは前極部(第23図)に徴小な臍状に突出している卵門丘の部分である。最も薄いのは卵門丘を取り巻く菊花状斑紋の周囲で、孵化の際に最初に喰破る個所はこの最も薄い部分に属している。

第23図 蚕卵直後の卵の縦断模式
A:前極、D:背側、P:後極、V:腹側。
a:前極原形質、c:卵殻、d:連絡管
m:卵門丘、n:卵核、p:周辺細胞質
v:卵黄膜、y:卵黄
外層の表面には無数の刻紋(卵紋)があるが、これは卵殻を分泌した包卵細胞の形のプリントされたものと考えられている(27,28,86,568,756)。刻紋の形は品種によって異なるから、蚕種の混同した場合などには判別の手がかりになる。卵門丘には外層を欠いている(29,960)。最内層は他の層と異なり、海綿状に近い構造のもののようである。卵殻の成分はキチンとかケラチンとか云う説もあったが、コリオニンである(567,719)。アミノ酸組成も調べられている(1057)。
卵門と気孔とが卵殻を内外に貫通している。卵門は前極部の卵門丘中央に開口している。1卵に1個であるが、開ロ部の直下で2-6本、普通3本の導管(精孔副枝)に分れて卵殻層内を通り(28,190,326)、その内端は長さ16-18μのラッパ状連絡管によって卵黄膜に連なっている。この連絡管は導管の延長であると云われているが(29)、導管末端部の形や、卵黄膜との接合部よりも導管との接合点で離れ易いことなどからみて、更に検討を要するように思われる。但し、卵黄膜のような膜状のものでないことは明かである。
卵門導管は、卵母細胞が7個の栄養細胞と連なっていた(Ⅰ2Aa)痕跡であるから、その数も7本であると云う説*があるが、少なくとも、 日本で普通にみる品種の卵には当てはまらない。桂(桂応祥 1934:広東特異蚕卵之形態与遺伝学的研究.仲愷農工学校研究報告 第2巻、第1号、1-260)の記載した広東産の蚕においても導管の数は3-4本が最も多く、1本、7本、8本のものは極く少ないと云う。
*朱洗・王高順・何家需・□詩宣 1954:家蚕混精雑交的研究(杉本行夫・亀井健三訳 1963:動物の多交配と遺伝性.新科学文献刊行会)
気孔は卵門丘を囲む菊花状斑紋部以外の卵殻全面に分布しているが(568)、刻紋との関係からみると、卵殻形成時に3個または4個の包卵細胞の接触点に当っていたと考えられる部分にのみ開口している(28,568,756)。従って、気孔の数は卵殻の分泌に関与した包卵細胞数のほぼ2倍に当る筈と考えられるが、大村・片岡(756)が5品種の卵について調べた気孔の数は1卵につき4661-9984個で、ほぼこの期待に一致すると云う。気孔の外面開ロ部の長径は3-4μであるが(30)、内方ほど細く、縦断面は漏斗状を呈する。
蚕卵を色素の水溶液に浸すと卵殻の中層および内層まで染色されるから(916,1070)、水溶液が卵殻層内に侵入することは確かであるが、最内層を通過して卵内にまで入るか否かは明かでなかった。 しかし、pH4.2の亜硝酸カリ(KN02)水溶液に蚕卵を浸すと極めて死亡率が高いので、これが郭内に容易に侵入することは確かである(709,1051)。ただ、亜硝酸カリの溶液に蔗糖を加えて浸透圧を高めると卵の生存率が高まったと云われているのは、2回の試験のうち1回だけがそのような結果で、他の1回は反対の結果を示していることからみてなお検討を要する。卵殻を貫通している気孔その他の細孔には空気が入っているので、浸透圧は働きにくい条件になっていると思われるが、実際に、濃厚な塩水に浸しても内容の収縮は起こらない。またN02が卵殻を通過したか否かをα-ナフチルアミンとスルファニル酸との混合液で定性してみると、亜硝酸イオンの卵殻通過は蔗糖の添加によっては影響を受けず、pHの影響の大きいことがわかった(未発表)。 pH5.6の溶液では蚕卵に対する亜硝酸カリの害がないと云われているが、この溶液の場合には亜硝酸イオンの卵殻通過をこの定性で確認することができなかった。なお蚕卵は、数カ月間水に浸しておいても、水を吸って膨潤し破裂すると云うようなことはない(Ⅵ3Bb)。
卵黄膜は卵母細胞の周辺細胞質から形成される無構造な膜である(496,501)。卵を生のまま生理食塩水中で解剖して漿膜に包まれた内容を取出すと、卵黄膜は卵殻内面に付着して残るが、卵を固定してから卵殻を除去すると卵黄膜は漿膜面に付着してとれてくる(未発表)。孵化の際には卵殻の内面に付着して残る(387a,983)。
卵黄膜の内部には原形質および卵黄が充満している。卵黄には脂肪性顆粒と蛋自性顆粒とが区別せられ(Ⅰ2Abii)、固定切片でみると、直径が前者は3.0μ(1.4-4.6μ)、後者は6.0μ(1.5-9.8μ)ぐらいである(290,496,501)。
卵黄膜の直下に卵黄顆粒を含まない細胞質の薄層(周辺細胞質)がある(555)。これは、受精後に分裂核が入って増殖、分化する部分で、胚発生上重要な意味を持っている。
卵門の付近で周辺細胞質の層の特別に厚くなった部分があり、前極細胞質と呼ばれることもあるが、産卵当時の卵核はこの部分に含まれている。交尾して順調に産み出された卵においては、産卵当時の卵核は卵母細胞の第一成熟分裂の後期の初めの状態であるが、未交尾蛾の産卵においては第一成熟分裂の中期であると云う(813-815)。
圧し潰しに対する蚕卵の硬さは主として卵殻の厚さによってきまる。硬さは卵の大小によってきまると云う報告もあるが、これは同一品種では、小さい卵が一般に卵殻も薄いためである。しかし、圧し潰しに対する硬さは、卵殻の一点の強さだけによるものではなく、卵全体としての構造が関係するから、小さい卵が必ずしも潰れ易いとは限らない(1,362,876,1079,1080)。高須(967)は、蚕卵の硬さは飼育時期、用桑その他の卵殻の厚さに影響すると考えられる諸条件によって相違するほか、催青の進むに連れて硬度が減じ、点青乃至催青期において最も潰れ易くなるが、孵化直前にはかなり回復することなどからみて、卵殻ばかりではなく卵内容物質の変化とも密接な関係があるものと考えている。
2 発生
A 受精
卵門から精子の侵入するのは卵が母体をはなれる数秒前である(Ⅰ2Ba)。精子の侵入によって第一に変化するのは周辺細胞質である。堅い卵殻で覆われている蚕卵においては、ウニなどにおけるようなこまかい観察や実験を行なうことはできないが、少なくとも周辺細胞質の表層に変化が起こって(ゲル化であろう)、表層と卵黄膜とが離れ易くなることは確かである(387a)。末交尾蛾の卵に比べて、交尾蛾の卵においては産卵当時の成熟分裂の進んでいるのも、精子侵入の刺戟によるものと考えられる。
精子の尾部は非常に長く、卵の長さの半分ぐらいもあるが(Ⅰ3Ab)、これが全部卵内に入ってしまうのか、精子の頭部が卵の周辺細胞質に達した処で尾部から離れるのかは確認されていない。盛んに産卵している交尾蛾を生理食塩水中で手早く解剖し、丁度、膣前庭にある卵を取出してみると、精子の長い尾部が卵門から出ているのを観察することができるが、前庭より下った位置の卵ではこれをみることができない。尾部が侵入してしまうのか、粘液腺からの膠着物が付与されるためにこれと一緒になってしまうのかは不明である。
1個の卵に侵入する精子の数は1個より多いのが普通で、11個入っていた例も報告されている(325,759,814,1074)。朱洗ら(前節参照)は、卵門導管が7本であるから、侵入する粒子数は最高7個であると云っているが、導管数によって制限は受けないもののようである。
1卵内に多数の精子が入っても、卵核と結合し得るのは普通の場合は1個の精子だけであるが、母親遺伝をするモザイク系統の雌蛾に、目標になる優性遺伝子ホモの雄蛾と劣性遺伝子ホモの雄蛾とを二重交尾させてみると、1個だけではなく他の精子も受精力を保持していることがわかる。母親遺伝をするモザイク系統の卵においては、正常の雌性前核のほかに1個の極体も別の精核と結合して受精する。しかし、こうしてできた分裂核は統合して1個体を作るので、二つの受精に由来する違った性質が体の部分に別々に発現して、モザイクや雌雄モザイク蚕になるのである(1046)。
このような試験結果から興味のあるのは、二重交尾をさせた2頭の雄蛾の精子は、交尾の順序に従って使用されるのではなく、混合して1卵内に侵入し得ることである。これは普通の系統の蚕においても同様で、1頭の雌蛾に優性遺伝子ホモの雄蛾と劣性遺伝子ホモの雄蛾とを二重または三重に交尾させてみても、初めに交尾させた雄蛾の子が特別に多いとは限らないから、受精嚢に入った精子は交尾の前後に拘らず混合使用されるものと考えられる。従って、このような交雑の場合に、交尾の順序を変えても優性の子供と劣性の子供との割合が同じ方向に片寄るようであれば、両雄蛾の精子の間に受精競争が行なわれ、受精力の強い精子が多くの卵を受精させたものと考えなければならない(1002,1003,1007,1008)。大村(753)によれば、幼虫期に出血させたり、蛹または蛾で長く冷蔵した雄蛾の精子は受精競走において無処理のものに劣ると云う。受精競走はその結果からみて選択受精とも云う。小村(708)は二重交尾をさせた場合における、両雄蛾それぞれの性質を受けた卵の産卵順序を調べている。
卵内に入った精子の核は卵核と結合するばかりでなく、条件によっては精核同志で結合して胚またはその部分を作ることがある。橋本(176)は、交尾蛾の産んだ産卵後1時間以内の卵に36.5-41.5℃の高温を1時間作用させると、精核同志の結合によって生じたものと考えなければ遺伝学的に説明し得ない個体や、モザイク蚕の現われることを報告したが、田島はこの現象を2精子メロゴニーと呼び、多くの実験を行なった(1046)。但し、メロゴニーと云う言葉がついていると本来のメロゴニー(卵片発生)とまぎらわしいので、最近では雄核発生(androgenesis)と呼ぶことが多い。
高温処理による精核同志の結合は細胞学的にも確められている(816)。交尾蛾の産んだ卵を高温で処理すると単為発生よりも雄核発生の起こり易いのは、精核に比べて卵核が高温や低温に対して弱いことが原因と考えられるが(181-183,819)、産卵後3時間目頃に処理卵の前極部付近をみると、卵の長径の1/5ぐらいの楕円圏内において精核が活発に運動しているから、これも精核の結合を前けるのであろうと云う(1030)。 雄核発生は低温処理(1013,1014)、炭酸ガス処理(1050,1052,1053)、などによっても起こる。
未交尾蛾の産卵に種々な刺戟を与えて単為発生を行なわせる実験(327-329,813,815-817,821)、蛾または化蛾に近付いた蛹の卵巣内から取出した卵の温湯処理による単為発生(70,179,180,204,210,211,896、897)などに関しても多くの報告がある。卵巣から取出した卵を処理して得られる単為発生蚕は全部雄である。これは第一成熟分裂において染色体が両極に分かれないためと考えられる(70,817)。交尾蛾の卵は産卵直後に処理しても、卵核だけが単独で発生することは極めて少ないと云う(181)。
未交尾蛾の、特別な刺戟を与えない自然のままの産卵においても、胚が単為発生によって発育を始めることは珍らしくない。不受精卵と云われているものを切片にして調べると、漿膜の不完全に形成されているもの、異常な胚盤形成の認められるもの、胚ができて死んでいるものなど、種々な発育階程の卵の含まれていることが観察される(828,1074)。これは人為単為発生の成功しなかった卵についても同様である(812)。ただ、自然単為発生においては孵化にまで達するものが極めて少ない(327)。
佐藤(813,814)によれば、精子の侵入した卵においては、80゚F(26.6℃)の場合には産卵後5分内外で第一成熟分裂が完了し、第二成熟分裂は産卵後60分頃から始まって80分頃に終り、雌雄両前核は産卵後2時間目頃に合体(受精)して第一分裂核を作る。第一分裂核はその後1時間以内に最初の分裂を起こすと云う。
これに対し、田島ら(1052,1053)は、適当な卵色遺伝子を持たせた雌雄を交配し、産卵後、0-20分の卵から始め、以後20分毎に一定時間の炭酸ガス処理を施こし、雄核発生その他の例外型および死卵の発現を調査した結果から、第一成熟分裂、第二成熟分裂、分裂核の第1回および第2回目の分裂完了時を、それぞれ、産卵後50、110、190および250分頃と推定した。村上・鬼丸(601)が蚕卵を圧し潰して細胞学的に調べた結果では、分裂核の分裂週期は40-50分と考えられたが、その後、X線による蚕卵致死率の変動から村上(600,602)が推定した分裂核の分裂核週期は60分である。大槻・村上(774a)の最近の報告によれば、第一成熟分裂は産卵後40分頃に終了し、分裂核の分裂は、少なくとも第3回までは同調で、周期は60分であった。
朱ら(Ⅵ1)は、二化性種の越年性卵においては、極体が卵膜(受精模であると云うが詳細不明)の外に放出されることが多く、極体を膜外に放出した卵は不越年性になるから、この放出が越年、不越年の目印になると云い、浸酸処理はこの極体を人為的に放出させて、不越年にする方法であると述べているが、極体の形成されるのは上記のように産卵後間もない時期であり、浸酸処理を行なうのは普通にはこれよりも約20時間後であるから、極体の追跡は困難になっている筈である(814)。
B 胚の発有階程
胚発生の知識が産業に直接結び付いていると云う点では蚕は他に比類のない動物と云えるが、蚕糸と云う立場だけから考えられたためか、最も得易い材料を極めて多数の人が手がけている割には、正しい意味での昆虫発生学の材料として用いられることの少なかったのも蚕である。
高見・北沢(935)は、蚕種保護の実用のために、蚕の胚発育を次ぎの30段階に分けている。
┌Stage 1 受精期
│Stage 2 分割期
│Stage 3 胚分化期
前休眠期 ─┤Stage 4 卵黄分割期
│Stage 5 だるま形期
│Stage 6 こけし形期
└Stage 7 へら形期
┌Stage 8 休眠Ⅰ期
休眠期 ─┴Stage 9 休眠Ⅱ期
┌Stage 10 前甲期
│Stage 11 甲期
越冬期 ─┤Stage 12 乙A期
└Stage 13 乙B期
┌Stage 14 丙A期
臨界期 ─┴Stage 15 丙B期
┌Stage 16 丁A期
│Stage 17 丁B期
│Stage 18 上唇突起発現期
│Stage 19 短縮期
│Stage 20 頭胸分節期
器官形成期─┤Stage 21 反転期
│Stage 22 反転完了期
│Stage 23 毛瘤発生期
│Stage 24 剛毛発生期
└Stage 25 気管螺旋糸発生期
┌Stage 26 点青Ⅰ期
│Stage 27 点青Ⅱ期
完成期* ─┤Stage 28 催青Ⅰ期
│Stage 29 催青Ⅱ期
└Stage 30 孵化期
*原著においては気管形成期に入れていた。
このうち、蚕種保護のために特に必要な階程だけを、特徴を強調して描いたのが第24図である。
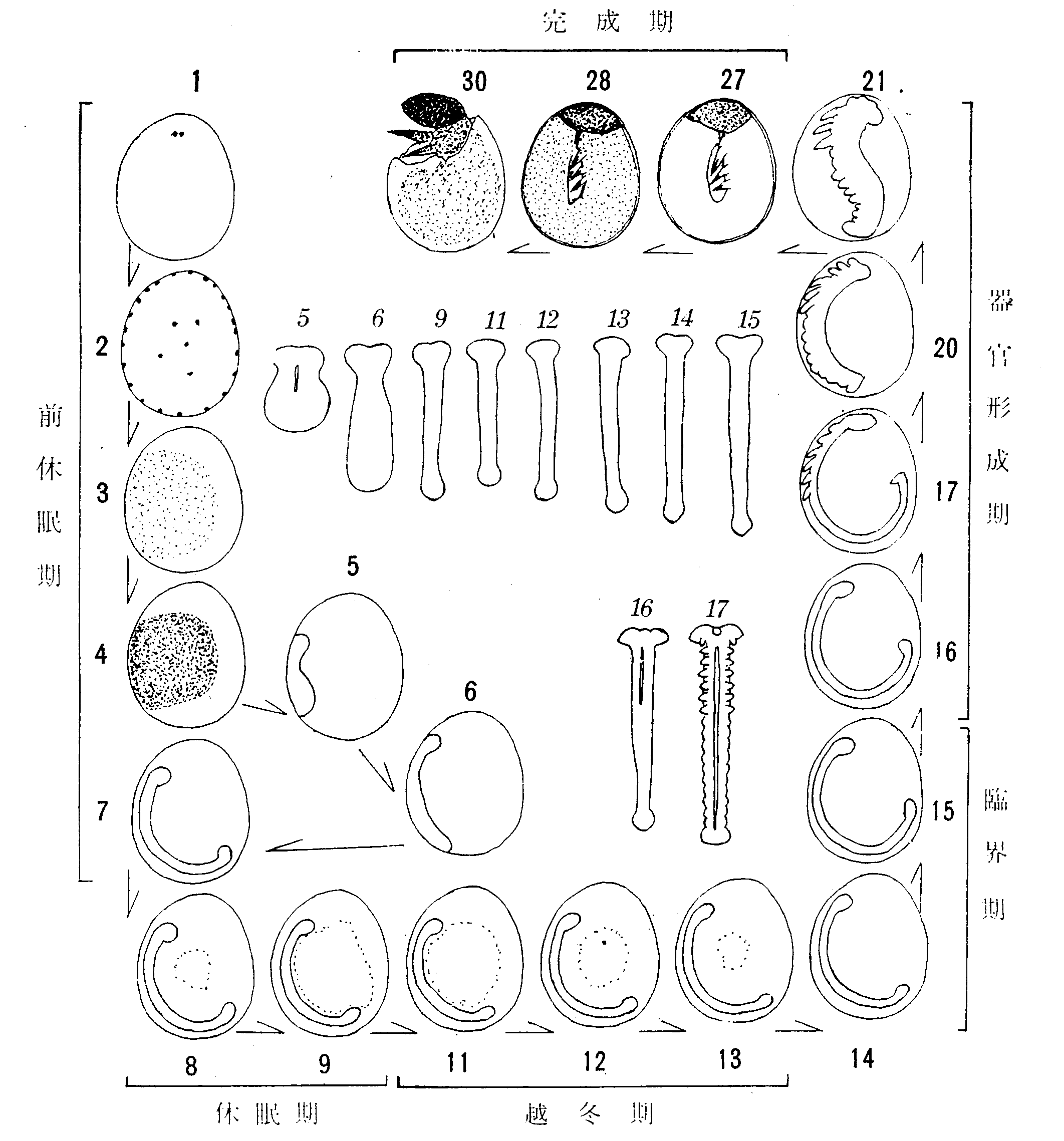
第24図 越年性卵の胚発生模式
ゴチックの数字はその発生段階(本文参照)の卵の縦断または透視。
イタリックの数字はその段階の胚を取出して腹面からみた輪郭である。
以下これによって解説する。発生経過は23-25℃における産卵後の時間である。
a 前休眠期
Stage 1 受精期 第二成熟分裂を終えた卵核(雌性前核)は卵内のほぼ黒点で示した位置で精核(雄性前核)と合体する。2時間目頃である。
Stage 2 分割期 受精によって生じた第一分裂核は3時間目頃から分裂を始め、以後、娘核は分裂を繰返えしながら周辺部に向って移動し、10時間目頃に周辺細胞質中に入り込み、12時間目頃にはこに1層の細胞層(胚盤)を作る(289,291,560,561)。一部の分裂核は卵黄細胞となって卵内に残る。
分裂核の分裂週期を1時間とすると、同調分裂が続けば8-9回の分裂で胚盤期に入る訳で、EphestiaやDrosophilaなどで知られているのとほぼ同じであるが、蚕卵においては何時まで同調分裂が続くのか正確にはわかっていない。
Stage 3 胚分化期 15-20時間の時期である。 15時間目頃から腹側を中心にして胚盤の広い範囲が厚みを増し、この部分が胚に発育するから、後に胚になる部分(胚域)と漿膜や羊膜に発達する部分(胚外域)とが識別できるようになる。識別できるようになった胚域は胚帯と呼ばれている。
蚕の卵は、一見、偏平な楕円体であるが、相称形ではなく、第23図に示すように尖りめの部分(前極)、その反対側の後極、膨らみのある腹側、その反対側の背側が識別できるが、胚は前極に頭部を向け、腹側に腹面を向けて作られるのであって(911)、正常な発生においてはこの向きが変わることは決してない。そればかりではなく、胚域のどの部分が、将来、胚のどの部分になるかと云うことも、胚域がはっきりしてくる以前の胚盤期、または分裂核が周辺細胞質に到着するより前から、既にかなりの程度に決定している(918,919,922)。宮が卵の種々な部分を焼き殺して生殖腺の発生を調べたのは(Ⅰ1)、この前決定の性質(484)を利用した研究である。
雌雄によって、催青卵期の卵の中での胚の右向き、左向きが違うと云う説があるが(ⅣEc)、卵の中で胚の形成されるのが腹側にきまっていること、産み出される途中の卵の廻転によってこの腹側が右になるか左になるかが偶然にきまり、反転によって胚の左右位が形成のときとは反対になることなどを考えれば、この説には理論上からも根拠のないことがわかる。
Stage 4 卵黄分割期 胚帯は腹側に向って、左右から(図で云えば表裏から)収縮してますます明瞭となり、20時間目頃には腹側を中心にして馬に鞍をかけたような形になる。
卵黄は、この頃までは蛋自性顆粒と脂肪性顆粒との単なる集積に過ぎないが、やがて卵黄細胞を含んだ小塊に切崩され初める(289,291,947,1157)。これを卵黄分割と云うが、卵黄分割は卵の周辺部から始まって次第に中央部におよび、胚の直下にも進行する。分割の結果、卵黄は卵黄細胞の含有物の観を呈するようになるが、この分割体(卵黄細胞)は卵黄粒団と呼ばれることもある。卵黄分割が完了すると、胚に卵黄が粘着しなくなるため、生で解剖した場合に胚が離れ易くなる。越年性卵の卵黄細胞においては脂肪性顆粒が核の周囲に集積しており、不越年性卵においてはこれが細胞内蛋白顆粒の間に分散している(926)。
この間に胚外域の細胞が胚帯表面を覆うように延びて漿膜と羊膜とを作る(238,904,921,1073)。漿膜細胞の大きさおよび数は品種によって異なり、母親遺伝をすると云う(333)。足立(15)は、産卵後40時間になっても、なお卵内から分裂核が卵の表層に到着して、既に着色した漿膜の形成に参加するように述べているが、そのようなことはない。
加温即時浸酸の適期はこの時期(20時間目)である。
この頃までの胚の発育は越年性卵でも不越年卵でも大差はないが、その後急に違いが現われてくる。外見に現われる越年性卵の最も著しい変化は、1日余りで漿膜細胞に色素形成が始まり、卵が淡紅褐色に色付いてくることである(332,1132)。卵の酸素消費量もこの頃から急に低下し(Ⅵ3B)、グリコーゲンの含量が減少する(Ⅴ2B)。
Stage 5 だるま形期 胚帯収縮が更に進むと左右から縊れが入り、胚の形に頭部と尾部との区別ができてくる。“だるま”形とはその形から名付けられたのであって、越年性卵においては約30時間、不越年性卵においては約25時間の頃である。
Stage 6 こけし形期 越年性卵の35時間目頃に当る。20時間目(Stage 4)に加温即時浸酸を施した卵は、条件によっても異なるが、浸酸後約18時間でこの時期になる。
即時浸酸種の冷蔵は18時間後に行なうのがよいとし、俗にダルマ期冷蔵と云われているが、即浸後18時間の胚は、だるま形よりも長くなった“こけし”形の時期であって(913,917,933)、真のだるま形期(Stage 5)に冷蔵すると成績が悪い(Ⅷ2Cb)。
完成した漿膜の表面には、普通気付かれないでいるが、薄い漿膜クチクラ(serosal cuticle)がある。これは36-48時間目頃に漿膜細胞から分泌される(556)。漿膜クチクラの形成と空気や液体の透過性との関係はわかっていない。
Stage 7 ヘら期形 越年性卵の3日目である。以後、休眠中を通じて、胚の外形は殆ど変らない。3日までは、胚を構成する細胞に分裂中のものがあるが、4日になると分裂はみられなくなる(386)。従って、越年性卵の胚は産卵後約3日で前休眠期の成長を完了するものと考えられる。
卵色は日に日に濃くなり、1週間ぐらいでその品種固有の色に落付く。卵内の3・ヒドロキシキヌレニンはこの色素形成のために消費されて、産卵後1日ぐらいから急激に減少するが、5-8日で減少が殆ど停止する(244,1132)。越年性の完成が不良な場合にはこの消費が少なく、また即時浸酸種においては越年性卵に比べて消費が少ない(355,357)。卵色は漿漠の色と卵殻の色、ときには透けてみえる卵黄の色もこれに加わった複合色である。越年性卵でも漿膜の着色しない突然変異型がある。
b 休眠期
水野の休眠期(Ⅵ2Bc)とは違う。何時からが休眠期であるのか、はっきりした区切りはできないが、2週間ぐらいで、一応、休眠と云われる状態になる(Ⅴ1Ba)。休眠Ⅰと休眠Ⅱも明確な区切りをつけることはできない。
Stage 8 休眠Ⅰ期 休眠期の胚の形は品種によってかなり相違する。前休眠期との相違で著しいのは胚の形よりも卵黄の状態の変化である。生体で解剖してみると、前休眠期には卵黄細胞が互に遊離しており、胚に付着しているものも離れ易いが、次第に離れにくくなる。同時に、中央部にある卵黄細胞が周辺部に向って移動勤し、集塊となって胚および漿膜に固着するため、卵の中央部に卵黄のない部分ができてくる(289,291,928,946,1078,1157)。この部分は次第に大きくなり、透過光線で透けて明るくみえるようになる。
Stage 9 休眠Ⅱ期 卵の中央の卵黄を欠く部分はますます拡大し、卵黄に堅く包まれた胚は、生体解剖では卵黄との見分けが困難な状態になる。
越年性卵はこのような状態で発育休止のまま夏秋期を経過する。卵黄の色は体液の色によって異なり、黄血蚕の卵は産卵当時から濃い黄色であるが、これとは違い、色素を形成して次第に着色するものがある。第2白卵(w2)種の卵が産卵後次第に橙色に着色するのは、漿膜にも若干色素ができるが、主として卵黄の着色によるものである。カンボージュなどの多化蚕の卵の着色にも同様のことがみられる。赤卵(re)の卵黄も橙色に着色する(652)。詳細に調べると、正常卵色の卵においても、卵内に点々と紅色の色素粒を形成しているものが珍らしくない(385)。
c 越冬期
自然湿度で保護してある蚕種の胚には、東京地方では12月初句になると、ぼつぼつと細胞分裂が現われる。しかし、その数は少なく、これを25℃で催青しても1ヵ月では孵化のみられないことが珍らしくない(386)。蚕の越年性卵は完全な休眠状態ではなく、このように一部休眠の醒め初めた状態で冬を迎えるのであって、低温に接触しなければ揃って発育するようにならないので、三浦はこの時期を対寒的発育期と呼んでいる(546,552)。
Stage 10 前甲期 催青すると胚が発育するから、休眠は脱しているが、完全に活性化した甲期とは違い、孵化までに非常に長い日数を要し、また発育が不揃いである。漿膜に堅くはり付いていた胚や卵黄が幾分ゆるんではいるが、形態的に休眠期とはっきり区別のできる特徴はない。第24図には省略した。
Stage 11 甲期 水野(577,580)の甲胚子に当る。甲胚子を水野が“所謂休眠期の時である”と記載しているために、今日の言葉での休眠と混同して、甲胚子は休眠胚であると誤解されていることがある。しかし、水野の云う休眠期は“休晩期に入りたる胚子は其後温暖に逢うことによって発育を始むるものである”と述べていること、および“甲胚子を摂氏17度半(華氏63度半)に1日間接触すれば凡そ乙A胚子を得る”と記述していることからわかるように、未だ発育を始めてはいないが胚が十分に活性化して、完全な発育を開始し得る状態に達した時期を指すものである。今日では越年性卵の胚が発育休止状態になることを休眠に入ると云うが、水野はこの時期に対しては休眠と云う言葉を使っていない。言葉は同じでも、当時の休眠期(水野以外にも同様の記載がある)は今日の休眠解消期に当るから、注意しなければならない。
また、水野の発生段階においては、甲胚子には中胚葉塊の環節的配列が認められない、と解説されているが、家蚕の胚には、前休眠期の終りから休眠期、越冬期、臨界期を通じて、中胚葉の環節状の配列が認められない時期はない。ただ、甲の頃は胚が短縮し、中胚葉塊が互に密接していて認めにくいだけである。この時期の中胚葉塊の形は品種によって異なる。どの部分のものがきまって大きいと云うこともない。
この時期の胚の形として、頭部の巾が広く、尾部ほど細くなって、頭葉と顎部との間に縊れのない図の示されていることが多いが、品種や標本の作り方によっても異なり、常にこのような形とは限らない。結局、完全に活性化はしているが未だ長さに伸びの認められない時期が甲と云うことになる。
生体解剖で観察すると、一部の卵黄細胞が遊離しているが、未だ無定形な集塊状を呈するものが多い。
Stage 12 乙A期 水野の乙Aに当る。甲より幾分長くなり、中胚葉塊相互の間隔がやや開き、胸節のものが他の部分のものに比べて幾分大きいが、明確な区別点はない。遊離の卵黄細胞が増加するが、これも連続的な変化に過ぎない。
Stage 13 乙B期 水野の乙Bに当る。胸節の中胚葉塊が他の部分のものに比べてはっきり大きくなるが、未だ体巾一杯には拡がっていない。卵黄細胞は大部分遊離するが、外形のすっりきりしないものが多く、大塊を作っているものもある。乙Aとの違いは連続的である。頭葉が大きくなり、体長も長いが、量的な違いで、乙Aとの聞の明確な区別点にはならない。
越冬期の胚には、暖い日が続くと細胞分裂の認められることがあるが、5℃以下の温度に保護されているものにおいては細胞分裂はみられない。この時期の胚発育は、休眠による透過性の変化と冬期の低温とによって脱水状態になっていた胚および卵黄細胞が、活性化に伴なって水分を取りもどし、伸びだした状態と考えられる(386,945)。甲は透通性が回復して伸び得る状態になったとき、乙Aはこれが伸び初めたとき、乙Bは伸長の終りに近い時期とみるべきもの(Ⅵ3Da)であるから、基準を設けてこれらの間をはっきり区切ることはむずかしい。蚕種保護の上からも、これらの時期の胚を形態によって区別することに苦心するよりは、胚が完全に活性化して、温度を上げれば順調に発育し得る状態になったときを確認することが大切である。水野が、17.5℃に1日おけば甲が几そ乙Aになり、翌日乙B、翌々日に丙A、更に1日で丙Bになると記載していることは、この意味で非常に含みのあることと考見なければならない。
d 臨界期
東京付近の自然温度においては、2月末から3月初めにかけてみられる越年種の状態である。頭葉が左右に開き、越冬期には体の中心線に沿って塊状に近い形で並んでいた中胚葉が左右に拡がり、18個の体節がかぞえられる。卵黄細胞は完全に遊離して卵内一様に分布し、卵の中央部の、透過光線で透けてみえた部分がなくなる。
越冬期から引続き蚕種を5℃に保護しておくと、胚はその温度の下で臨界期までは発育し、100日ぐらいおけば細胞分裂も現われるが、2.5℃では活性化はするが250日たっても臨界期の形態を示すに至らなかった(945)。
三浦(546,552)は、寒冷に接触させる必要がなくなり、温度の上昇に連れて胚が発育する状態になったとして、この時期以後を対暖的発育期と呼んでいる。
Stage 14 丙A期 水野の丙Aに当る。頭葉が開いて大きく、体は伸びて細長な感じになる。特に体後半の伸長が著しい。中胚葉塊は左右に拡がって帯状を呈する。卵黄細胞は完全に遊離する。
Stage 15 丙B期 水野の丙Bに当る。頭葉は十分に開き、倒三角形を呈する。胸節の中胚葉塊は左右に分れる直前で、先端で付着し合ったハの字形を呈する場合が多い。尾節は後端が尖り菱形に近くなる。水野は胸部の1、2節に微かんに正中線の現われたときまでを丙Bとしているが、本書においては正中線(神経溝である)の現われたものはStage 16 とする。
細胞分裂を伴なわずに伸長し得る極限の階程で、これ以後の発育は細胞分裂を件わなければ進行しない(386)。休眠期および越冬期の胚を生体で生理食塩水中に取出すと、著しい屈曲、捻転をあらわすが、この時期になると屈曲は全く起こらない。胚の吸水伸長の完了したことを示すものであろう。
水野は丙Bを胚の発育経過中で最長の時期であるとして最長期胚子と名付けたが、実際にはこの時期よりも次ぎの丁A、丁B期の胚の方が長いから、梅谷(1101)は水野の丙Bを細長期、水野の丁Aを最長期と呼んでいる。岩崎は丙Bを臨界期と呼んでいる。呼び名が混乱しているから、古い文献を読む際には注意が必要であるが、普通には、最長期と云えば丙B期のことである。
蚕種の保護、取扱いは胚の発育階程を基礎にして行なうと云われているが、その発育階程のうちで最も重要なのはこの丙Bの時期でる。単式冷蔵においても中間手入れ方式の冷蔵においても、この時期まで胚を発育させて冷蔵するのが越年種の冬期保護の常法である。この時期を過ぎると冷蔵に耐える力が低下するから、鑑定を誤まらないように注意しなければならならない。上に述べたように、越冬期の胚の鑑定には形態的に明確な基準が乏しいが、越冬期の胚を調べるのは、丙Bの鑑定を誤まらないための補助的な調査であると云ってもよいぐらいで、蚕種保護上大切なのは丙Bである。
中胚葉が左右に拡がって、胸節においては、ハの宇の上端が付着し合ったような形になるが、それが未だ左右に分れない時期が丙Bで、体の正中線に神経溝がみえ初め(胸節から始まる)たものは既にこの時期を過ぎている。胚の形は品種によっても異なるから、大きさとか、長さとか、頭葉の形とかなどのように変異の多いものよりも、15℃で保護した場合に、神経溝のみえ初める1日前、付属肢の現われ初める2日前と云うことを基準にして、丙Bの形に慣れ
ることが必要である。
ただ注意しなければならないのは、丙B期の胚が2.5℃では100日、5℃では60日の冷蔵に耐えると云われているのは、活性化と胚の発育とが調和して進行している場合には、胚が丙Bになったときの生理状態が冷蔵に適していると云う意味であって、条件が違えばこの形態と生理状態との間にずれが生じ、丙Bが必らずしも冷蔵適期を示さないことに注意する必要がある(Ⅶ1Da)。丙B期が重要であると云うのは、丙Bと云う形態を通して推定される生理状態が重要なのであって、形態が重要なのではない。
e 器官形成期
胚に各種の器官の形成される時期である。
Stage 16 丁A期 胚の正中線に神経溝が現われ、頭葉が角ばってくる。水野の丁Aの時期である。上に述べたように、梅谷はこの時期を最長期と名付けたが、実際には顎節および胸節に付属肢の生じた頃(丁B)の胚の方が長い。もっとも、梅谷のF胚子(水野の丁Aに相当すると云う)には“上顎、下顋、下唇の3対の突起および胸脚の3対の突起徴かに認め得”られると云うから(1101)、水野の丁Aよりは進んだ時期であって、ここにもまた混乱がある。
Stage 17 丁B期 顎節および胸節に付属肢が発生する。続いて腹節にも突起が現われる。水野の丁Bに当る。胚は反転期以前では最も長いが、胚発育期間を通じて云えば、最も長いのは孵化直前の完成胚であるから、最長期と云う名称はこの時期に対しても適当ではない。
不越年性卵の胚は産卵後2日でこの階程になる。
Stage 18 上唇突起発現期 上唇突起は丁A期から発生が始まるが、これが次第に前方に向って移動し、2個の突起となって頭端に現われるのがこの時期である。この期の初めには尾節の巾が腹部後端の巾より広いが、次第に狭ばまり、後期には両者の巾が等しくなる。ほぼ水野の戊Aに相当すると考えられる。第24図には省略した。不越年性卵は産卵後2日半でこの時期になる。
Stage 19 短縮期 体長目立って短縮、尾節が腹部の後端より狭く、小さくなる。気門および絹糸腺陥入。ほぼ水野の戊Bに相当すると考えられる。不越年卵は3日でこの時期になる。第24図には省略。
Stage 20 頭胸分節期 体長は著しく短縮する。顎節が次第に集合し、将来の頭部になる部分と胸部になる部分ととの区別がつくようになる。触肢突起が前進して口陥と同列になる。尾節は非常に小さい。ほぼ水野の戊Cに当る。不越年性卵の3日半。
Stage 21 反転期 これまで卵周に沿って卵の腹側に位置を占め、背面に湾曲していた胚に湾曲転換が起こり、同時に胚が背側へ転位する運動の全期間で、ほぼ水野の己Aに当る。不越年性卵の4-5日目である。ヘロルド腺発生(668)。
中田(668)は、第1腹節から後方の体節が卵周を離れて内部に移動し始めた時期を反転前期第1期、第1腹節から後方が殆ど一直線になった時期を反転前期第2期、第7腹節から後方が腹面湾曲に転じた時期を反転後期第1期(第24図、第25図a)、第1腹節から後方が脱脂湾曲に転じた時期を反転後期第2期(第25図b)に細別している。
反転の機構については諸説がある(60,145,268,484,920,1100)。
反転すると直ぐ胚脱皮が起こる(703)。脱皮は引き続いて2回行なわれる(未発表)。前胸腺の働きとは関係がない(951)。
Stage 22 反転完了期 湾曲転換が完了し、口部の諸器官はロ陥の周囲に集まり、ほぼ蟻蚕にみられるような配置をとる(第25図c)。不越年性卵の5日目。
Stage 23 毛瘤発生期 毛瘤発生。尾角が生じる。ほぼ水野の己Bに該当すると思われる(第25図d)。不越年性卵の6日目。この頃から胚に脈博が認められる(1197)。
Stege 24 剛毛発生期 剛毛発生。胸肢に爪が生じ、大顎に鋸歯形成。単眼の着色始まる。図省略。不越年性卵の6日半。
Stage 25 気管螺旋糸発生期 気管に螺旋糸発生、大顎着色(第25図e)。不越年性卵の7日目。

第25図 反転後期から点青初期までの胚(中田)(668)
a:Stage21 反転後期第1期、b:Stage21 反転後期第2期
c:Stage22 反転完了期、d:Stage23 毛瘤発生期
e:Stage25 気管螺旋糸発生期、f:Stage26 点青Ⅰ期
f 完成期
胚体の完成する時期である。生体で解剖してみると、取出した胚は生理食塩水中で活発に体を動かす。
Stage 26 点青Ⅰ期 胚は未だ漿膜を飲み込んでいない。漿膜があるために卵の外部からはみえないが、内部では胚の頭部表面がキチン化し、褐色を呈する。臍孔は完全に閉鎖。固定した胚を色素で染めると、胴部が未だよく染まる。ほぼ水野の己Cに当るものと思われる(第25図f)。不越年性卵の7日半。気管にガスが入る(637)。
Stage 27 点青Ⅱ期 胚が漿膜を飲み込み、着色した頭部は卵殻を通して青みを帯びてみえる。点青または目付きと呼ばれる時期で、孵化の前々日に当る。胴部は未だ着色が極く淡いが、色素に染まり難い。孵化を揃えるために、この時期から光を遮断して催青するのが普通である。不越年性卵の8日目。
Stage 28 催青Ⅰ期 胚は頭部ばかりでなく胴部も着色するため、卵全体が青みを帯びた色にみえる。催青卵または青み卵と呼ばれる時期であるが、胴部の着色が未だ十分に濃くないため、生卵をそのままみると、頭部の色が濃く、胴部との識別が容易である。この段階は点青後期と呼ばれていることもある(986)。不越年卵では9日目。
Stage 29 催青Ⅱ期 胴部の着色が濃くなり、生体のままの卵の外観では、卵全体が一様に青み、頭部をはっきりと識別し難い。孵化直前の状態である。不越年性卵は9日半でこの時期に達する。催青卵の卵殻を切破って胚を出し、乾燥を防いで飼育すると、自然に孵化したものと同様に発育し得る。室賀(613)は欧19号の青み卵になったばかりの卵から胚を取出して発育させることができたと云っているが、大村・菅生(757)によれば、孵化前日20時の胚は取出しても正常と大差なく育ち、同12時の胚は表皮柔かく扱いにくいが15%が上蔟し、同3時の胚は柔かく水々しているが3%が3令に達した(以後の飼育中止)。孵化前々日9時の胚は白く水々しているが、4%が3令に達した。
Stage 30 孵化期 孵化。不越年性卵では10日目。孵化の行動については室賀(607)、森山(595)、中島(653)、竹内(983,984)らの観察がある。
不越年性卵や人工孵化種の胚は、Stage7 から直接Stage14に眺んだような形態変化を極く短時間のうちに経過し、卵黄細胞の移動も漿膜の着色も起こらない。ただ不越年性卵が越年性卵と異なるのは、胚に18体節の全部が現われないうちに、頭胸部になる部分の体節構造が特に明瞭になり、正中線も認められることである(129,450,668)。産卵後約10日で孵化するが、器官形成期に入ってからの経過は、時間的にも形態的にも、越年性卵と大差がない(第154-156表)。
3 生理
蚕種の取扱いに関係の深い問題だけについて述べる。
A 水分
産卵当時の蚕卵は平均65%ぐらいの水分を含んでいるが、日のたつに連れてその絶対量が減少し、6月採種の越年性卵は翌年3月までに約6%の水分を失なう(671)。しかし、この間に固形分も4-5%減少するので、水分率としては殆ど変化がないとされている。勿論これは保護条件によって相違し、湿度の低い場合には水分の減少が大きい(Ⅵ3Db)。催青に着手すると蚕卵重量の減少は著しく、孵化前日までに約10%減るが、水分率はほぼ66%を保持している(367)。蚕卵は、卵殻の構造上、長時間水に浸しておいても内部へ水を通さず、水分率に殆ど変化がない(783)。
このように、程度の差はあっても、水分は産卵後減少の一路を辿り、タト部から補給されることはないのであるが、卵殻の着いたままの卵全体ではなく、毛細管で採種した卵の内容だけについて測定すると、胚発育の時期に応じて著しい水分率の増加または減少が認められると云う(466,467)。黒田らはこの水分微量測定法によって調べた結果、外部からの吸水の考えれない蚕卵の中で、産卵後3日間に6%以上も水分率の増加していることに注目し、この増加した水は、胚の成長に伴なって貯蔵養物が“酸化分解され又縮合される等新陳代謝の結果生ずる”代謝水であると考えた(467)。
梅谷(1101,1106)は、この方法によって調べると、越年卵においては、1)産卵後2日目(ダルマ期)、2)7-10日目(アレイ期)、3)越冬後、胚が発育を始めて付属肢突起を形成する時期、および4)反転後に水分率の低下がみられるが(第26図)、これらは何れも代謝作用の緩慢な時期であって、低温に耐える力も強いとし、広く卵態越冬の昆虫をみると、総べて蚕のこれらの時期の何れかに相当する段階で越冬しており、蚕の胚は2)の時期で越冬すると論じている。但し、この解釈はバッタの卵の生理作用の消長と一致しない点があるが、それは測定法の相違によるのではないかと云う。伊藤(278)は、催青中の卵において、梅谷が抵抗力の弱いと云っている時期にほぼ該当する反転前乃至反転期および孵化前に尿酸量の減少することを認め、抵抗力との平行関係に注目しているが、生理的な意義は不明で、平行現象として指摘したに過ぎない。

第26図 越年性卵における水分量の時期的変化(梅谷)(1106)
越年性卵においては、休眠が進むと卵黄細胞が周辺部に移勤し、卵の中央部に液質の部分ができ、冬中この状態が続くのであるが、これが水分微量測定の値にどのように影響するかについては言及されていない。しかしこの卵黄細胞の偏在は凍結実験の結果には次ぎのように影響している。
青木(46)によれば、越年性卵を-36℃ぐらいの低温で凍結させて凍結曲線を調べると、-15~-20℃で過冷却が破れて凍り初め、温度は一且-5℃付近まで急上昇するが、再び徐々に下がる。この凍結曲線の下降部は産卵後1日目には滑かで、凍結点は一つだけであるが、3-4日目頃から下降曲線の-13℃付近に第2凍結点が現われ初め、10日目には極めて明瞭な平坦部として認められる。このような凍結曲線の形は産卵後100日間も続くが、多価アルコールの存在(Ⅴ2B)とは関係がない。凍結した卵を速かに融かして再凍結を行なうと、1度目に完全に凍結させた卵には第2凍結点が現われないが、1度目の凍結を第2凍結点の現われる前に止めて融かした卵にはこれが現われること、未受精卵の凍結曲線にも第2凍結点は現われないこと、などから考えると、休眠の進行に伴なって卵内に凍り易い部分と凍りにくい部分とのできることが考えられる。これは卵黄細胞の移動によって卵の中央部(この部分に熱電対を挿入して測定した)に液質の部分のできることとよく平行している。越冬後、丙B期頃に達した卵においては第2凍結点が現われないとも聞いている。
越年性卵にグリセロールおよびソルビトールの増加するのは冬期間の凍結防止に役立っているのではないか、と云う考えはこの凍結実験の結果によって否定されたが、蚕卵は休眠状態で冬を越すものではなく、これらの多価アルコールの減少し初めたか、もしくは減少してしまった状態で厳冬期を過すことからみても、その生態的意味を大きく考えることはできない。
蚕卵からの水分蒸散は、即時浸酸種についての測定によれば(698)、25℃、湿度75%の場合に、浸酸後3時間半で最高(1.470mg/g.hr)を示し、以後急激に減少し、翌日には0.551、3日目には0.519で最低に達する。その後は次第に増加し、9日目(孵化前日)には初日にほぼ等しい価(1.313)に回復する。この変化は梅谷の再発育期以後の代謝水の増減(第26図)と傾向を同じくしていると云う。
休眠に伴なって胚および卵黄細胞から水が出て、それらの組識は収縮しているのであろうと云うことを先きに述べたが、この時期の蛋自性卵黄顆粒には中性赤による顆粒状染色(コアセルヴェーショソと考えられる)が起こりにくいことからみて、蛋白質分子の水和性が強くなっているのではないかと云う想像もできる(929,948)。
B 呼吸
a 不越年性卵
催青中の毎日の酸素消費量(105,364,680,681,683,684)、同じく炭酸ガス排出量(368,893)などには多くの研究がある。
不越年性卵および浸酸卵の炭酸ガス排出量または酸素消費量は、催青初期には日々の増加が明瞭でないが、反転後から増加し、点青期頃に急増する(第97表)。
| 卵 令 | 不越年種の炭酸ガス排出量 (五大州、26℃)(g) |
即時浸酸種の酸素消費量 (日111号、25℃)(g) |
| 第2日 3 4 5 6 7 8 9 10 |
8.7417 10.0207 12.3540 8.4248 9.9410 14.8400 21.9980 31.4351 32.2519(一部孵化) |
8.072 7.248 9.083 7.438 9.514 12.065 13.803 16.605 31.977(催青卵) |
これらの排出量および消費量は温度によって著しく変化するが、西郷(781)によれば、点青期の酸素消費量と温度との関係はArrheniusの式

によって表わされ、μの値は5-15℃では25,000、15-25℃では15,700、25-37℃では6,000であり、Q10は5-15℃で4、15-25℃で2.5、25-35℃では1.5であると云う。
川瀬(364)は、催青中のチトクロームcオキシダーゼ作用の増大が7日目(9日目に1部が孵化)までは顕著でないがその後急増することと、卵の酸素消費量が7日目以後に著しく増加することとの間には関係があるものと考えている。
入戸野・中曾根(684)は、休眠中の卵の酸素消費量は酸素分圧によって著しく影響を受けるがCOによる阻害を受けないこと、休眠を終えると酸素分圧の影響が減じるがCOによる阻害が現われ、殊に催青末期のCOによる阻害の著しいこと、COによる阻害は光線の有無によって影響されることなどから、休眠中の呼吸にはフラポプロテイン系酵素が、休眠を終えた催青中の卵の呼吸においてはチトクローム酸化酵素が、それぞれ末端酵素として働いているのであろうと考えた。
西沢ら(681)によれば、反転直後2日間ほどは青酸による酸素消費の阻害が特に著しい。
山ロ・清水(1165)は、卵そのまま、卵の磨砕物、胚だけを取出したもの、および胚の磨砕物の酸素消費量を比較し、催青中の胚においては、呼吸作用の70-40%は胚の“構造”に帰すべき部分、30-60%は酵素その他の、細胞の基礎的な生化学的組成に帰すべき部分であろうと云っているが、詳細は不明である。
催青中を通じて、呼吸商が0.6-0.7であることから(368,683)、そのエネルギー源は主として脂肪であろうと考えられている。
催青中の酸素分圧は170mmHg内外が最適で、200または140mmHgでは催青死卵が出ると云う(604)。
催青中の所要空気量あるいは換気回数などを決定する基礎資料としては、前記鈴木の炭酸ガス排出量が用いられている。重量で出ている排出量を体積に換算し、その量だけの炭酸ガスを許容量まで稀釈するにはどれだけの空気量が必要かと云う計算をするのであるが、これには気圧、温度、空気中の炭酸ガス量、炭酸ガス許容量などを測定しなければならない。
簡便法としては、炭酸ガス許容量(P)を1%(1㍑中10cc)、空気中の炭酸ガス量(Q)を0.04%(1㍑中0.4cc)とし、気圧は無視して計算する方法が用いられている(605,606)。体積に換算した炭酸ガス総排出量をK(cc)とすれば、所要空気量(Y)は
![]()
である。室賀(620)は催青中の所要空気量を、この方法によって蚕種10g当り88.2㍑と計算した。許容量を0.5%にとった鈴木(884)の計算では178㍑となっている。
しかし、これらの値は、蚕種を実際に催青した結果から考えられる所要空気量に比べて、非常に大きな空気量である。室賀自身も、毎日1回換気すれば、10gの蚕種に対して1日2㍑でほぼ足りると云い、三浦(547)は、40,000粒の蚕種の催青には、1㍑の容器に収容して1日2-3回換気すればよいと記載している。古い横田ら(1194)の成績によれば、容績1㍑のガラス器内に密封して、全期開催青しても害のない蚕種(不越年)の量は2-3蛾(1蛾400粒内外)以内であるが、1㍑に15-16蛾分を密閉しても、催青の前半だけならば被害がないと云う。杉山・高見(877)のガラス器に密封した試験結果では、10gの蚕種は全催青期間を通じ20㍑の空気があれば十分に孵化した。1日2㍑の空気量では、催青初期には十分であるが、末期にはやや不十分であった。芦田(68)は、即時浸酸種1gを30日間冷蔵(5℃)する所要空気量を20cc以上としている。
人工孵化種を催青初期に輸送する場合には、輸送に2日かかるものとすれば、蚕種10g当り多めにみて12㍑の空気量があれば、密閉して送っても呼吸に支障がないと云う計算(591)が示されている。これは上記の方法によって計算したものであると云うが、実際問題としては、このように大きな容積の荷造りをして蚕種を送ることはない。横田ら(1194)によれば、木箱であれば、越年種、不越年種ともに1㍑に100蛾分を密封しても催青初期8日間は被害がなく、この倍量を入れても初期5日間以内ならば差支えなかった(温度は75゚F前後)。
このような計算と実際との不一致を、室賀(622)は、計算量は呼吸障害を蒙るような空気には絶対に接触させない場合を考えた量であり、実験例として報告されているのは、呼吸障害を蒙るような不良な空気に接触しても、その期間が短かいために害がなかった場合であろうと考えているが、なお検計すべき問題である。
卵殻だけを除去した卵内容全体を液体培地で培養すると、点青期まではよく発育して、漿膜も嚥下するが、ここで気管系を通じての呼吸に切り変わるため(637)、胚は溺死する(941)。
b 越年性卵
越年性卵が休眠に入ると呼吸量が著しく減少するが、これは単なる量の減少ではなく、青酸やCOによる阻害の減少から考えてもわかるように質的な変化である。越年性卵における酸素消費の青酸阻害は産卵後2日目頃までが最も著しく、1.5×10-4Mで阻害が現われるが、4日目以後では3×10-4Mになり、以後90日になってもこの状態が続くと云う(785)。
休眠に伴なう糖代謝の機構を追究した茅野(Ⅴ2B)の研究は、休眠卵の呼吸について注目すべき結果を示している。グリコーゲンからのソルビトールおよびグリセロールの生成反応には多量のNADH2およびNADPH2が必要である。不越年性卵においては、これらの補酵素はミトコンドリアの呼吸酵素系によって速かに酸化されるが、越年性卵においては、正常の呼吸酵素系が何処かの点で遮断されるために、これらの補酵素の酸化が止まり、多価アルコールの生成に向けられるようになる。休眠に入ってからの脂質の減少量は、低下した酸素消費量(第27図)によっては到底CO2およびH2Oに完全に酸化し得ないほど多量であるから、脂質からの水素はそのまま酸素に行くのではなく、何らかの中間代謝物に移されているものと考えなければならないが、これは呼吸商の増大からも推定される。

第27図 休眠初期の蚕卵における酸素消費量および呼吸商(Chino)(82)
○:酸素消費量、×:呼吸商。測定は25℃。
NADH2およびNADPH2は解糖、ペントース燐酸径路およびリンゴ酸酵素によるリンゴ酸塩の酸化などの過程で作られて多価アルコールの生成に用いられ、この生成は無酸素状態でも阻害されない。しかし、休眠を終えて多価アルコールからグリコーゲンの再合成される反応には酸素が必要である。
更に特異なのは、蚕卵ホモジネートの吸収スペクトルによって確認できるチトクロームはb5であり、cは、孵化の3日ぐらい前になって微量に存在するのではないかと考えられる程度の量しか見出されないことである。チトクロームcオキシダーゼの存在はスペクトルで確認できる。b5はミトコンドリア分劃には極く僅かで、主として、脂質に富む分劃に含まれており、この分劃にはチトクロームcオキシダーゼも多い。チトクロームb5およびチトクロームcオキシダーゼの最大吸収部は、休眠に入ると次第に消失し、産卵後10日ぐらいで殆ど認められなくなる。休眠から醒めると再び現われるが、休眠初期における多価アルコールの出現はこれらの消失よりも早く、また休眠から醒めると低温下でもグリコーゲン量は回復するが、チトクロームb5およびチトクロームcオキシダーゼは催青を始めなければ出現しないから、チトクロームの消失が多能アルコールの生成の第一義的な要因とは考えられないと云う。
茅野は実験の結果を綜合して、前休眠期および休眠後の催青初期には、NADH2またはNADPH2からフラボプロテイン、チトクロームb5、チトクロームc、チトクロームcオキシダーゼを通る特殊な径路で酸素への電子伝達が行なわれ、催青末期に筋肉が発生するようになるとミトコンドリア分劃にチトクローム酵素が集中して、普通の電子伝達系が働くようになるのではないかと考えている。石川(258)は休眠に伴なう上記補酵素の酸化型と還元型との消長を調べている。
越年卵を冬季間、自然温度の水に浸漬しておくと、胚に付属肢の形成が始まる前までは、発育はややおくれるが自然温度で保護した無浸漬卵に比べて孵化歩合は劣らず、寧ろ高い場合があるが(782)、付属肢形成が始まってからも浸漬を続けると、急激に孵化歩合が低下すると云う。また、産卵後3カ月ぐらい高温で保護した卵を5℃の水に浸すと230-250日の浸漬に耐え、高温30-60日の卵は同じく140-170日の浸漬に耐えたと報告されているが、著者らの試験(末発表)では、産卵後、25℃、30日で浸水した場合に、孵化歩合が50%に低下する浸漬日数はこれに比べて非常に短かく、5℃で30日、25℃で3日に過ぎなかった。その原因は明かでない。著者らの場合には、水の腐敗を防ぐために卵を消毒し、滅菌蒸溜水に浸漬した。
C 発育
a 胚の成長
熱湯で固定した越年卵から胚を取出し、その乾物量を測定した山口ら(1154,1162)の結果によれば、
催青期間の10-20%を経過した最長期乃至突起発生期
〃 45% 〃 反転期
〃 70% 〃 気管形成期あるいは口器着色期
をそれぞれ中心とする成長率の高い時期があり、30%(反転前)および50-60%(反転後)を中心にする成長率の低い時期がある。木下(380)もほぼ同様な結果を報告している。
山口らはこの成長率の変化は梅谷の考え(Ⅵ3A)によく一致すると云うが、別の見方をすれば、最長期乃至突起発生期は盛んな細胞分裂と共に器官形成の始まる時期であるから成長率の高いのは当然であろうが、反転期を中心とする時期の重量成長には、背面形成によって体内に包み込む卵黄の重量がみかけの成長として加算されているのではなかろうか。また、気管形成期あるいは口器着色期と云うのが、木下の報告のように点青期のことであるとすると、漿膜を飲み込んだための体重増加が関係しているかも知れない。これらの点を吟味した上で検討する必要がある。
| 測定月日 | 体 長 (μ) |
体 巾 | 胚の呼び名 | ||
| 頭葉(μ) | 胸部(μ) | 尾節(μ) | |||
| 1月15日 30 2月 5日 25 3月 5日 10 15 20 25 30 4月 5日 10 |
1,305 1,286 1,165 1,252 1,332 1,431 1,403 1,564 1,435 1,546 1,454 1,779 |
344 320 315 350 347 351 352 393 351 347 338 399 |
220 198 202 229 207 220 203 246 207 214 199 210 |
225 192 196 204 187 181 194 213 189 218 194 216 |
┐ │ ├水野甲、梅谷A-B │ ┘ ┐ ┴水野乙A-乙B、梅谷C ┐ │ ├水野丙A、梅谷D ┘ 水野丙B、梅谷E(細長期) |
| 固定時期 | 体長 (mm) |
体 巾(mm) | 胚の発育階程 | |||
| 第4環節 | 第6環節 | 第8環節 | 第13環節付近 | |||
| 1月15日 30 2月15日 28 3月15日 30 |
1.45 1.53 1.75 1.75 2.11 2.19 |
0.19 0.19 0.21 0.23 0.20 0.22 |
0.20 0.20 0.24 0.25 0.21 0.23 |
0.17 0.19 0.21 0.22 0.19 0.21 |
0.17 0.18 0.21 0.21 0.18 0.20 |
甲 乙A+ 乙B- 乙B+、丙A 丙B 丙B+ |
b 卵重の変化
蚕卵の重量は産卵後の経過に連れて減少する(第100表)。
| 蚕品種 | 蚕種の種類 | 基準卵重 | 催 青 | 孵化 | 毛蚕 | |||||||||
| 直 前 | 第1日 | 第2日 | 第3日 | 第4日 | 第5日 | 第6日 | 第7日 | 第8日 | 第9日 | |||||
日122号×支115号 支115号×日122号 |
春採越年 〃 |
g 0.2772 0.2822 |
% 8.0 3.9 |
% 8.6 4.2 |
% 8.8 4.4 |
% 9.1 4.5 |
% 9.4 4.8 |
% 10.3 5.5 |
% 11.4 6.6 |
% 12.8 8.3 |
% 14.6 10.9 |
% 16.4 ─→ |
% 18.1 12.5 |
% 29.1 28.7 |
|
日122号×支115号 支115号×日122号 |
秋採越年 〃 |
0.2637 0.2629 |
3.8 2.4 |
4.1 2.6 |
4.2 2.7 |
4.4 3.0 |
4.9 3.3 |
5.7 4.1 |
7.1 5.4 |
8.9 7.2 |
11.4 9.9 |
─→ ─→ |
14.2 11.3 |
27.6 26.2 |
|
日122号×支115号 支115号×日122号 |
春採即浸 〃 |
0.2953 0.2886 |
─→ ─→ |
1.2 1.2 |
2.2 2.2 |
2.8 3.3 |
3.9 4.4 |
5.2 5.5 |
6.6 8.0 |
10.2 ── |
── ── |
─→ ─→ |
12.2 10.3 |
26.2 25.6 |
|
日122号×支115号 支115号×日122号 |
春採即浸冷 〃 |
0.2981 0.2890 |
─→ ─→ |
2.9 3.6 |
4.0 4.9 |
5.2 6.2 |
6.3 7.6 |
8.5 10.3 |
11.8 ── |
── ── |
── ── |
─→ ─→ |
14.3 13.3 |
26.8 29.5 |
|
日122号×支115号 支115号×日122号 |
春採冷浸 〃 |
0.2965 0.2952 |
─→ ─→ |
0.8 0.6 |
1.3 0.9 |
1.7 1.2 |
2.4 1.7 |
3.3 2.6 |
4.7 3.7 |
8.0 6.2 |
── 9.1 |
─→ ─→ |
11.4 11.0 |
28.4 28.3 |
山口(1158)は、催青中の卵重量の減少は直線的ではなく、初期はやや緩慢であり、催青期間の50-60%を過ぎると急降下すると云い、水分率や卵汁の理学的性状(浸透圧、電気伝導度、比重、粘性)などの変化をも調べた結果を綜合して、卵の発育には反転期頃を分岐点とする生理的二期があるとし、反転期は生化学的作用が旺盛であると云う梅谷の説(Ⅵ3A)とはいささか矛盾すると考えている。この場合、反転期の中心は、24℃内外の催青において、丙Bから蟻蚕完成(孵化の前日)までの経過の大約45%の点であった。
鈴木の測定した不越年性卵の炭酸ガス排出量(第97表)をみると、産卵後5日目頃に一時的な排出量の減少が認められるが、徳永(1056)はこの現象に注目し、鈴木の成績においては五大洲の卵期間の44%、青熟(表には省略)の同40%の時期にこの排出量の低下していること、また水野ら(574)の調査した即時浸酸種の減耗量についても、日本種春採り3品種平均卵期間の41.7%、同秋採り3品種平均および日支交雑種2品種平均のそれぞれ卵期間44.4%の時期に減耗量割合が一時的に低下していることを指摘し、不越年性卵の成長にも越年性卵にみられる休眠に類似の現象があり、そのために二期的な成長をするのではないかと考えている。しかし、炭酸ガス排出量については兎も角として(川瀬ら(368)の4日目の低下は鈴木の場合のように顕著ではない)、卵重量の減耗は、山ロの場合、水野らの場合および第100表の成績がそれぞれに異なるようであるから、なお検討の余地があるように思われる。即浸卵の4日目頃に減耗量の低下するのは、布目らのみている蒸散量の低下(Ⅵ3A)と一致するのかも知れない。
D 環境
a 温度
催青中の胚発育は温度によって著しく影響を受ける(第101表)。
| 蚕 品 種 | 20℃区(%) | 24℃区(%) | 28℃区 |
| 日 1 号 支 4 号 支 7 号 支 9 号 欧 7 号 欧 9 号 日107号 正 白 支101号 新 白 |
89 92 84 81 82 88 94 90 95 96 |
90 89 90 93 86 90 93 91 95 95 |
85 93 83 93 69 88 89 84 93 95 |
松村・石坂(515)が第101表と同様な3回の試験結果を平均した孵化歩合は、20℃区92.54%、24℃区93.02%、28℃区88.88%であったが、20℃区は24℃区または28℃区に比べて孵化の不斉な場合が多かった。
10品種平均の胚発育速度(催青日数の逆数)は、湿度72%場合に、20℃区0.064、24℃区0.090、28℃区0.106であった。孵化した蟻蚕の体重は28℃区が最も軽く、絶食生命時間は 24℃区>20℃区>28℃区であった。
赤卵や白卵の越年種には孵化の悪いものが多いが、これらは夏期の保護温度および催青温度を25℃よりも低目にすると孵化がよくなる(964,1115)。
これらの結果を綜合すると、胚の発育に最適な催青温度は20℃以上25℃以下であることがわかる。
なお蟻蚕の体色は催青温度の高いほど淡く、低いほど黒味を帯びる。蟻蚕の体色に温度の影響するのは胚の形態が完成し、皮膚の着色する時期である(1135)。松村(512)および長島(635)はメラニン色素の形成を調べ、温度と蟻蚕体色との関係を化学的にも証明した。松村は、産卵直後の保護温度が高いと卵色の淡いこと(Ⅴ2A)をも同様にメラニン色素の濃淡によるものとしているが、これは主として3・ヒドロキシキヌレニンの色素形成のための消費量が温度によって相違するためであろう。蚕卵においてはメラニン系色素は検出できないと云われている(717)。褐円斑紋の発現は、反転期の催青温度が25℃以下の間は温度の上昇に連れて低下するが、25℃以上になると反対に高まるらしい。即ち10℃≧15℃>20℃>25℃<30℃<35℃であった。
越年性卵の漿膜細胞色素粒は、産卵後、高温の続く間は細胞内に一様に分布しており、そのために卵色は濃くみえるが、気温が低下すると核が細胞の一端に偏在するようになり、色素粒もその周辺に集まってしまうので、卵色が淡くみえるようになる。冬が過ぎて気温が上ると色素粒は再び細胞内に一様に分散する(533,534,670,1137)。自然温度で蚕種の冬期保護を行なう場合に、漿膜色素の分散状態が卵の活性比状態を知る一つの指標になるように云われることがあるが(954)、特定な品種を特定な地方で毎年みているような場合は別として、一般的には、色素数の移動と卵の活性化状態とは必らずしも一致せず、色素粒の集積したときに既に活性化のかなり進んでいるものもあれば、全く活性化していないものもある。胚の形とこの移動との間にも直接の関係はない(1087)。
休眼中の温度については既に述べた(Ⅴ1B)。
休眠を終えた胚が孵化まで発育し得る最低温度は12.5℃であるが、点青期以後ならば10℃、催青卵ならば7.5℃でも孵化し得ると云われている(905)。若い胚ほど低温においてもよく発育するのに(第103表)このような結果になるのは、低温においては発育に非常な長期間を要するために、若い胚から初めると途中で活力が低下してしまい、孵化にまで達し得ないのであろうと考えられているが、臨界期までの発育は胚の伸長に過ぎず(Ⅵ2B)、物質合成を必要とする器官形成期の成長は低温では十分に進行し得ないのであろうと云うこと(702)も考える必要がある。
胚の長さの伸長を発育とみるならば、休眠を終えたばかりの胚は0℃においても僅かながら発育する(第28図)。

第28図 冷蔵温度別の胚発育(高見・戸谷ら)(945)
日124×支124のF2卵(晩秋採り)。12月3日に洗落し5℃、2.5℃および0℃に分けて冷蔵した。
最上段の数字は月日で、その時測定した胚の長さの分布を縦線の長さで表わす。短かい横線は平均値。
核分裂の認められなかった場合は細い縦線、認められた場合は太い縦線で示した。縦線の下の数字は
催青2週間孵化歩合。点線の場合には胚長は測定しなかったが、孵化歩合は調べた。
この図において注目されるのは、活性化、胚の発育および核分裂はそれぞれに必要な最低温度を異にしていることである。
0℃においては活性比はするが、胚の発育は僅かに進むだけであり、核分裂は全然起こらない。
2.5℃におては、活性化および胚の発育(丙Bまで)は進むが、調査した期間内では核分裂は超らなかった。5℃においては、活性化、胚発育および核分裂が共に進行する。即ち、臨界期までの発育は伸長を主とするものであり、物質合成を伴なう真の成長はその後に始まると考えられる。
この点に関連して興味があるのは鈴木(893)の炭酸ガス排出量に関する試験である。9月3日産卵の青熟越年卵を2月12日から151日間、+5℃、-2.5℃および-5℃に保護して、その間の炭酸ガス排出量を測定した結果から蚕卵1kg当りの1日量を計算すると、+5℃ 0.28660g、-2.5℃ 0.00379g
、-5℃ 0.00322gで、-2.5℃と-5℃との差は小さいが、+5℃と-5℃との間には生理的に大きな断層のあることがわかる。
水野によれば、冷蔵中の胚発育は第102表、胚の階程別発育速度は第103表の通りである。
| 冷蔵当時 の胚 |
冷蔵温度 (℃) |
冷蔵日数 | ||||||||
| 20日 | 40日 | 60日 | 80日 | 100日 | 120日 | 140日 | 160日 | 200日 | ||
| 甲 | -2.5 0 2.5 5 7.5 |
甲 甲 甲 (-)乙B (-)丙A |
甲 甲 甲 (+)乙B 丁B |
甲 甲 甲 (-)丙A (+)戊 |
甲 甲 甲 丙A 戊B |
甲 甲 乙B 丙B (+)戊B |
甲 (+)甲 乙B (-)丁A (+)戊B |
甲 (+)甲 乙B (-)丁A (+)戊B |
甲 乙A 乙B (-)丁A - |
甲 - - - - |
| 丙B | 0 2.5 5 7.5 |
丙B 丙B 丙B (-)丁B |
丙B (+)丙B (+)丙B 丁B |
丙B (+)丙B 丁A 戊B |
丙B (+)丙B (+)丁A (-)戊C |
丙B (+)丙B (-)丁B (-)戊C |
丙B (+)丙B (-)丁B (-)戊C |
丙B (+)丙B (-)丁B (-)戊C |
丙B (+)丙B (-)丁B - |
- - - - |
| 目的の 発育期間 |
保護温度 (℃) |
発育所要日数 (日・時間) |
積算温度 (時間) |
目的の発生段階に達した胚の割合 |
| A 休眠期(甲) から 最長期(丙B) |
30.0 27.5 25.0 22.5 20.0 17.5 15.0 12.5 10.0 |
3・12 3・12 3・12 3・12 3・20 4・10 4・17 6・10 11・03 |
2,520 2,310 2,100 1,890 1,840 1,855 1,695 1,925 2,670 |
5%。他は総べて若く、極めて不斉。 8%。他は大部分若く、極めて不斉。 24%。 〃 〃 44%。不斉。 55%。やや不斉。 90%。斉一。 92%。 〃 90%。 〃 91%。 〃 |
| B 最長期(丙B) から 反転期 |
30.0 27.5 25.0 22.5 20.0 17.5 15.0 12.5 10.0 |
3・14 3・14 3・14 4・02 5・02 7・13 9・19 14・16 33・17 |
2,150 1,935 1,720 1,715 2,130 2,263 2,350 2,640 4,045 |
全部。斉。 〃 〃 〃 〃 〃 〃 93%。他は若く、ほぼ斉一。 75%。他は老幼やや不斉。 67%がほぼ反転、他は老幼やや不斉。 52% 〃 他は老幼不斉。 57% 〃 〃 |
| C 反転期 から 催青期 |
30.0 27.5 25.0 22.5 20.0 17.5 15.0 12.5 10.0 |
4・06 4・10 4・18 5・18 7・16 11・20 16・00 30・00 55・00 |
2,550 2,385 2,280 2,415 2,760 3,550 3,840 5,400 6,600 |
斉一。 〃 〃 〃 〃 〃 〃 〃 〃 |
第103表において注目されるのは、Aのように若い区間の発育所要日数は、30℃でも15℃でも著しい相違がないが、Cのように進んだ区間の発育所要日数は温度が低いと非常に長いことである。これを逆に云えば、胚の発育程度の揃っていない蚕種を高温で催青すると発育の開きがますます大きくなるが、低温で催青すると、進んだ階程の胚の発育は比較的おそくなり、おくれた階程のものの発育が比較的進んで、全体としての発育の開きが小さくなる。これは蚕種の取扱い上実際に利用される重要なことがらである。
低温のために発育を抑えられていた胚は、高温に移すと発育が非常に速い。例えば、第28図の場合に、2.5℃区4月6日の胚は平均体長1.43mmであったが、これを10℃に移して2日おいたものは1.58mm、15℃1日では1.55mmに発育しただけで核分裂は認められなかった。これに対し、25℃に移したものは1日で2.01mmとなり全部の個体(11頭)に核分裂が認められた。
蚕卵の耐え得る低温の限度についての奥村(707)の試験によれば、越年卵を2月に-20℃の恒温に冷蔵すると、2日間でも著しく孵化歩合が低下したが、北海道の冬季間のように、-20℃以下の日はあっても、間歇的な場合には被害がなかった。仲野(656)によれば、休眠期胚子(甲胚子の意味)は-5℃に14日接触させても孵化が斉一であったが、-10℃14日間または-15℃3日間では孵化不斉になった。
丙B以後、孵化前日までの催青中の卵の耐寒性について、日9号×支108号、日114号×支114号および欧18号×支107号を用いて行なった山口(1156)の実験によれば(第104表)、催青期間12日の場合の5日目(経過歩合約42%、反転期)頃において、その前後に比べて耐寒性の弱い時があるように思われると云う。なお、この場合の不発生卵は大部分が催青死卵で、普通死卵は1-5%(10%を越える例は極めて少ない)であったから、この程度の低温は胚に対して直接の致死作用をおよぼすものではないと考えられる。
| 催青日次 | 低温接触 | |||
| -10℃、6時間 | -20℃、6時間 | -10℃、24時間 | -20℃、24時間 | |
| 第 1日目 2 3 4 5 6 7 8 9 10 11 12 |
90% 81 83 75 64 80 74 81 67 14 10 0 |
74% 82 75 67 63 73 40 24 6 0 0 0 |
79% 82 72 80 78 80 70 23 2 0 0 0 |
78% 67 71 57 29 12 14 2 0 0 0 0 |
先きに述べた青木の凍結実験(Ⅵ3A)においては、-15℃~-20℃で凍結が始まったが、これは熱電対の挿入によってにじみ出た卵黄に氷ができ易いためで、自然状態で凍結の始まる温度はこれよりも低いと云う。
梅谷(1114,1116,1124)は交尾した雌蛾の産んだ卵を受精前に5℃と2.5℃とに冷蔵した処、5℃のものは60日間冷蔵した後にも70%以上の受精率を示したが、2.5℃においては1日間の冷蔵によっても受精率が30%ぐらいに低下したと云い、これは卵核ではなく精核が被害を受けるのであって、2.5℃と5℃との間に精子生存の臨界点があると考えた。これに対し、橋本(183)は、産卵後5-20分の卵を2.5℃、0℃、-2.5℃にそれぞれ42-43時間冷蔵した後に、高温処理を施して雄核発生(Ⅵ2A)を行なわせた結果から、産卵直後の低温に対しては精核よりも卵核が弱いと報告している。
滝沢・玉沢(1012)は、交尾蛾の産んだ1時間以内の蚕卵を冷蔵すると、-20℃においては2日間、-15℃においては10日間、-5℃および0℃では60日間ぐらいで受精力を失なうが、5℃においては60日間でも受精力を失なわず、しかも40日前後で淡褐色に着色し初め、受精卵と不受精卵との区別ができるようになるから、5℃では受精が行なわれると云っている。また、蛾を過冷却すると雄蛾よりも雌蛾の受精力の減退が著しいから、梅谷の考えには同意し得ないとしている。交尾蛾の卵を産卵後30分以内に冷蔵した結果からみると、-5℃と-10℃との間に抵抗力の臨界点があるらしい(1011)。
産卵後1-2時間目の卵が産卵直後の卵よりも低温(0℃)に対する抵抗力の弱いのは(618)、この時期に第二成熟分裂および受精の行なわれるためと考えられ、低温ばかりでなく、種々な障害に対して弱い時期である。山ロ(1155)は、産卵後25℃で2時間目を蚕卵標準抵抗力曲線の第1降下点と呼んでいる。第2降下点は産卵後12時間目の、胚盤から胚帯の形成され初める時で、人工孵化による畸形が生じ易い。20時間目以後は障害に強くなる。
蚕卵に対する低温の影響については、上記のように、条件の設定と受精または孵化とを直接に結び付けた報告が大部分で、その過程の分析がないために、結果の相違があってもその原因を究明し得ない場合が多いが、冷蔵によって卵内にある種のアミノ酸が増加すると云う報告のあることは注目される。
上原(1091)は、即浸冷蔵種の孵化の悪くなる前の、冷蔵期間25日頃から、卵内に遊離のプロリンが増加し、40-45日ではやや減少すること、ロイシン、フェニールアラニンも同様な動きをすることを報告し、高橋(914)もプロリン、グルタミン、グルタミン酸、ヒスチジンなどが冷蔵期間の長くなるのに連れて増加するのを認めた。高橋(915)は、また、冷蔵期間が長くなって障害の出初めた卵の磨砕液に熱水を加え、その遠心上清に1%のHgSO4またはHgCl2を滴下すると白色沈澱が生ずるが、 これは増加したプロリンその他のアミノ酸、あるいはそれに近い低分子の物質が沈澱するのであろうと考えた。
佐々木ら(809)は、蚕卵の遊離アミノ酸の研究中に、上記の現象に注目して実験を行なった。プロリン、ヒスチジンは、産卵直後(珍らしい取扱いである)から9日間冷蔵し、出庫当日浸酸した卵においても(浸酸直後)、冷蔵前に比べて著しく多かったが、プロリンはその後、反転期まで増加を続け、発蟻前に幾分減少し、ヒスチジンは発蟻まで増加を続けた。しかし、プロリンは浸酸および冷蔵を施さない卵*の反転期にも著しく多かった(催青著手前の測定は行なっていない)。プロリンソ、ヒスチジンの少ない産卵直後の卵の除蛋白濾液を加水分解すると、これらのアミノ酸が増加するから、産卵直後の卵にはプロリン、ヒスチジンなどを構成要素とする低分子のペプタイドまたはアミノ酸誘導体が存在し、卵内容の異常な組織分解によって、それらのアミノ酸の増加を来たすのであろうと云う。
(註:*1955年夏蚕用の秋花×銀嶺であると云うが、春採ならば浸酸しなければ反転期まで進まない筈であるし、
もし越年種ならば冬期の冷蔵を経ている筈であるから、この意味が不明である。)
化学の立場からではなく、蚕種の立場で問題点を生化学的に検討し、蚕種の取扱いとその結果とを結ぶ過程を明かにすることは今後心要な研究方向である。
催青中の卵を35℃に1日間接触させると2割程度の死卵が生じ、3日間の接触で殆ど全部が死卵になるから、蚕卵発育温度の最高限度は、実用的には35℃と考えられる(580)。
高温に対する抵抗力を胚の発育段階別にみると、小針らが(398)即浸種を40℃(湿度75%)に24時間接触させて孵化を調べた結果では、胚帯が卵表から離れて卵内に落込む頃(浸酸1時間後)、気管形成の始まった頃(浸酸当日を1日として数えた6日目)、頭部着色の頃(同7日目)および漿膜を飲み込む頃が弱く、中田(668)が不越年種を46℃-48℃(湿度20-35%)に20分および40分間接触させた結果では、分裂核分散、気門発生、反転(前期の第1期および第2期)および気管螺旋糸発生の各期が弱かった。越年種の催青中に45℃(湿度50-55%)に30分間接触させた木下(380)の成績では気門形成期、反転期および点青期が弱かった。反転の前後および点青期頃が共通的に弱い。
蚕種の抵抗力については、蚕種取扱いの実際問題として、以下の諸章において更に述べる。産卵直後の卵に対する高温の影響(Ⅳ5Aa)および休眠と保護温度との関係(Ⅴ1Ba)については既に述べた。
b 湿度
催青中の湿度が低いと孵化に悪形響がある。松村ら(514)の成績(第105表)によると、60%区は75%区および90%区に比べて明かに孵化歩合が劣っている。75%と90%との優劣は明かでなかった。7品種平均の催青日数は、25℃の場合に60%区11.33日、75%区10.98日、90%区11.00日で、75%および90%は60%に比べて短かかったが、75%と90%との差は明かでなかった。
| 蚕 品 種 | 催 青 湿 度 | ||
| 60% | 75% | 90% | |
| 日 1 号 支 4 号 欧 9 号 正 白 新 白 支4号×正白 新白×欧9号 |
78% 74 73 77 92 93 86 |
80% 78 89 83 95 92 88 |
83% 76 86 87 93 98 89 |
| 平 均 | 82 | 86 | 88 |
道家(88)によれば温度と湿度とは相関連して催青中の卵に影響する。総孵化歩合について云えば、95%以上の孵化歩合の得られる温湿度は15-28℃、50-95%の広範囲に亘るが、実用孵化歩合85%以上の得られる範囲は24-28℃、70-90%で、最適条件は25℃、85%であると云う。
孵化の際の湿度に関しては興味のある実験がある。室賀(613)は、最初の発蟻をみたときに卵面に桑葉を撒布すると、無撒布の対照区に比べて孵化が促進され、桑の乾葉を撒布しても同様の効果があるから、この場合の促進効果は湿度の影響ではなく、桑の香気の影響であるとした。これに対し、福田・橘田(118)は、さくら、いちょう、ぽぷら、のざわ菜などの葉を撒布しても桑と同様の効果がある;乾葉撒布では、室賀の結果とは違って効果が少ない;湿度を100%にした容器内においては、桑その他の生葉または乾葉を入れても殆ど効果がない;定湿にしたデシケーター中に1蛾の催青卵を分割して入れ、午後10時から翌朝10時までおいて孵化を調べると、第106表の結果が得られた、などのことから、上記の孵化促進効果は桑の香気によるものではなく、湿度の影響であろうと結論した。
| 蚕品種 | 湿 度 | |||
| 約17% | 50-60% | 70-80% | 100% | |
| 太 平 長 安 |
15% 18 |
16% 21 |
42% 46 |
71% 76 |
室賀・丸山(623)は、春採り越年性卵(欧7号および正白)を産卵後24時間目から湿度60%と80%との2区に分け、産卵後70日間は平均27.14℃(25℃以上30℃以下)、その後50日間は5℃、次いで0℃に25日間(冷蔵中の温度は両区同一とあるが、詳細の記載なし)保護した後、磨砕して生体含有水分率を比較した。生体含有水分率と云うのは卵殻を除いた部分の水分百分率であるが、5回の測定結果の平均は、正白80%区71.94±0.12%、60%区71.87±0.18%、差0.07±0.14%、欧7号80%区72.13±0.16%、60%区71.88±0.08%、差0.22±0.13%で、有意の差がなかった。蚕卵の生体重(対1,000粒)は10回平均が、正白80%区472.1±4.9mg、
60%区451.6±4.4mg、差20.5±1.2mg、 欧7号80%区634.6±4.9mg、
60%区618.8±5.1mg、差16.0±1.0mgで、何れも80%区の卵が重かった。(註:計算が合わない)。
産卵直後の多湿は蚕卵に対して有害で、特に温度の高い場合および低い場合にはその害が著しい(第107表)。
| 日 数 | 温度 | |||||
| 30℃ | 25℃ | 20℃ | ||||
| 湿度 | ||||||
| 75% | 98% | 75% | 98% | 75% | 98% | |
| 3日 5 10 20 |
97% 97 97 89 |
95% 94 87 56 |
94% 94 97 93 |
94% 91 76 41 |
95% 95 95 93 |
96% 95 91 58 |
山口・小林(1163)は、大シヤーレに硫酸、食塩水および水を入れて湿度0-100%の7区を作り、蚕種を入れた無蓋の小シヤーレをこの中に納めて越年種の冬期保護試験を行なった。供試したのは欧18号および改安の春採りで、10月23日から翌年4月10日までを上記の湿度区に分けて保護し、その間、毎月1回、卵の新鮮量、乾物量および孵化歩合を調べた。温度は自然温度である。
卵の新鮮量は何れの区においても、保護中に漸次減少し、減少の度合は低湿区ほど大きかった。乾物率はこれと反対に、低湿区では増加し、多湿区では減少の傾向を示した。
生物体にとっては、乾物量と水分量との一定の割合、即ち水分平衡を保つことが生理上望ましいものとするならば、乾物率の増減の最も少ないのが最もよい環境湿度と云うことになると想定したのであるが、このような湿度は、改安に対しては75%、欧18号に対しては50%であった。しかし、実際に孵化歩合の最もよかったのは90%区で、次ぎが75%および対照区で、最も悪かったのは、改安では0%区、欧18号では100%区であった(第108表)。
| 蚕品種 | 湿度 (%) |
孵化歩合((%) | |||||
| 11月 | 12月 | 1月 | 2月 | 3月 | 4月 | ||
| 改 安 | 0 10 30 50 75 90 100 対照 |
44 42 43 42 51 49 39 46 |
48 55 58 50 88 86 73 77 |
38 56 53 57 78 89 73 75 |
33 41 39 43 60 77 64 76 |
35 46 29 40 71 69 64 65 |
18 32 34 30 62 63 50 60 |
| 欧18号 | 0 10 30 50 75 90 100 対照 |
2 1 4 6 0 4 0 3 |
71 61 62 72 65 65 58 66 |
83 82 83 87 87 83 70 90 |
86 91 85 87 93 95 76 87 |
90 87 88 96 91 95 72 89 |
81 82 88 86 90 92 69 94 |
胚の発育は一般に多湿区が早く、また揃っていた(第109表)。
| 湿度 (%) |
蚕品種 | |||||
| 改安 | 欧18号 | |||||
| 調査時期 | ||||||
| 2月 | 3月 | 4月 | 2月 | 3月 | 4月 | |
| 0 10 30 50 75 90 100 対照 |
乙A-乙B 〃 〃 〃 〃 〃 〃 〃 |
乙A-乙B 〃 〃 〃 乙B 〃 〃 〃 |
乙B-丙A-丙B 〃 〃 〃 丙A-丙B 丙B 〃 〃 |
乙A-乙B 〃 〃 〃 〃 〃 〃 〃 |
乙A-乙B 〃 〃 〃 〃 乙B 〃 〃 |
乙A-乙B 〃 〃 乙B-丙A 丙A 〃 乙B-丙A 丙A |
試験全体を通じて、改安は区間の差が明瞭で、特に50%以下の乾燥に弱い傾向があり、欧18号は乾燥に強く、寧ろ100%のような多湿に弱い傾向を示した。この相違については、卵の大小、卵殻の厚さ、卵内容の相違なども考慮しなければならないとしている。
結論としては、冬期間(1-6℃)の蚕種保護の適湿は、催青の場合と同様に、75-90%であると云うのであるが、乾物率の増減を通して考えられる蚕種の代謝作用と保護環境との関係については論議が尽されていないように思われる。
なお、活性化の早さおよび全期間を通じての孵化歩合から考えて、改安は正常な状態の蚕種ではなかったように思われる。
催青温度と化性との関係についてはⅤ1Aa参照。
c 光線
渡辺(1148)によれば、点青期までの胚の発育は暗所におけるよりも明所において幾分速いが、その差は催青温度が20℃の場合に明かであって、25℃の場合には明瞭でない。点青期以後になるとこれとは反対に、明所におけるよりも暗所において胚の発育が進み、その差は低温よりも25℃以上の高温において明瞭である。更に発育が進んで孵化の段階になると、暗所のものは明所のものに比べて約9時間孵化がおくれる。これが、発育の進んだものの孵化を抑えて発育を揃え、一斉に発蟻させる暗催青法の基礎になっているのであるが、光に対する感受性は蚕の品種によって著しく相違している(421,429,954)。
恒明、恒暗および明暗交替(18時間毎)で催青した蚕種の孵化を比較すると、孵化は明暗交替区が最も早く、恒明区がこれに次ぎ、恒暗区が最もおくれるが、25℃恒明で催青した蚕種が若干孵化し初めたときから明暗の条件を種々に変えてみると、第110表のような成績が得られた。
| 試験区 | 光の条件 | 蚕品種 | |||||||||||
| S | N | ||||||||||||
| 経過時間 | |||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 1 | 2 | 3 | 4 | 5 | 6 | ||
| 1 2 3 4 5 6 7 |
150lux 恒明 暗・明(150lux)1時間交替 恒暗 5lux 恒明 5lux・30lux1時間交替 30lux 恒明 30lux・150lux1時間交替 |
151 0 0 12 15 218 179 |
162 138 0 31 243 151 159 |
31 3 0 27 23 48 20 |
38 146 0 17 48 50 61 |
36 2 0 13 27 30 27 |
35 89 0 32 63 41 64 |
94 69 66 99 79 103 101 |
54 41 59 41 38 19 46 |
33 29 12 46 32 30 39 |
23 36 8 27 15 17 25 |
15 14 6 12 13 15 14 |
10 24 8 17 12 13 14 |
これによってみると、光感受注の高い品種Sにおいては、恒暗にすると6時間の観察中、孵化が完全に抑えられた。5luxは恒暗と恒明との中間の作用をし、30luxでは150luxと殆ど同じ程度の効果があったが、150luxに比べれば明の効果が幾分弱いようである。Nは光感受性が非常に低い。
点青期以後を暗黒にしておき、若干孵化し初めたときから5、30、90、450luxの光を各1分間、5分間および450luxの光を60分間照射し、以後再び暗くして暗中での各区の孵化の遅速を比較しても(第111表)、NとSとの違いは明瞭である。
| 照射時間 | 蚕品種 | 調査項目 | 光の照度(lux) | 450lux 60分間 |
||||
| 0 | 5 | 30 | 90 | 450 | ||||
| 1分間 | N | 総孵化卵数(粒) 初め1時間の孵化歩合(%) 同上指数 |
65 66 100 |
67 67 102 |
58 81 122 |
69 65 98 |
49 57 86 |
56 83 127 |
| S | 総孵化卵数(粒) 初め1時間の孵化歩合(%) 同上指数 |
450 58 100 |
194 63 109 |
192 71 123 |
238 79 137 |
288 80 139 |
322 98 170 |
|
| 5分間 | N | 総孵化卵数(粒) 初め1時間の孵化歩合(%) 同上指数 |
271 60 100 |
202 70 117 |
160 61 103 |
247 65 109 |
154 64 108 |
148 75 126 |
| S | 総孵化卵数(粒) 初め1時間の孵化歩合(%) 同上指数 |
388 31 100 |
361 62 197 |
449 66 210 |
542 73 232 |
549 69 220 |
445 90 286 |
|
Sについてみれば、1分、5分ともに90luxと450luxとでは殆ど差がない。450lux・60分区の孵化の多いのをみると、照度だけではなく時間の影響することがわかる。孵化の誘発と云うことだけならば5lux・30秒でも効果は認められるが、実際問題としての孵化の促進には不十分である。しかし、発蟻抑制のための暗催青中にはこの程度の光でも抑制効果の妨げになる。本実験においては、調査のための蟻蚕の掃落しは写真用安全燈の下で手早く行なった。
以上のように、蚕卵の孵化は明暗によって著しく影響を受けるが、これとは別に、恒暗で催青してそのまま孵化させても、または恒明で催青したものを孵化前に恒暗にして暗中で孵化させても、一定の時間々隔で集中的に孵化の起こる現象がある。この孵化の週期(リズム)については多くの研究がある(985-994,1021-1027,1031)。
漿膜細胞の色素粒は低温によって集合し、温度の上昇によって分散するが(Ⅵ3Da)、光線の照射はこの分散を促進し、3・ヒドロキシキヌレニンからの漿膜色素の形成をも促進するもののようである。また、光は、点催青期の卵においては蟻蚕の斑紋色素の形成を促進し、発生初期には斑紋色素を減少させる傾向もあると云う(635)。
d その他の条件
点青期の蚕種を種々な気圧のもとにおくと、+100mm、-50mm、-100mmのような比較的小さな変化の場合には、常圧区よりも死卵歩合の減少する傾向が認められるが、その影響は処理時間6、24、48時の間では差がなかった(696)。また、点青初期の蚕卵を常圧より30mm低い気圧の下に20時間および40時間おくと常圧区に比べて、総孵化歩合に変りはないが、酸素消費量が増加し、孵化の促進がみられたと云う報告もある(1199)。
しかし、点青以前の蚕卵を-20mmで3日、5日、8日などの長期間処理すると催青日数が長くなり、孵化が劣るので、1蛾の卵を3分し、標高の違いによってこの程度の気圧差のある3カ所(東京、松本、小淵沢)に分けて催青試験を行なったが、孵化歩合に一定傾向の差が認められなかった(1198)。 但し、これらの試験においては、空気の組成などを調べてはいない。
長田(640)は0.5m/sec または1m/sec程度の気流があると催青日数が僅かながら短縮することを認めた(第112表)。しかし、孵化の際には妨げになるもののようで、卵殻に穴をあけたまま死んでいるものがやや多かった。
| 関係 湿度(%) |
気 流 |
催青4日17時間の胚 | 点青期に達した胚 | 催青日数 (日) |
孵化歩合 (%) |
|||||
| 調査卵 数(粒) |
反転前 のもの (%) |
反転中 のもの (%) |
反転後 のもの (%) |
調査卵 数(粒) |
催青8日 12時間に おいて(%) |
催青9日 12時間に おいて(%) |
||||
| 25 | 有 無 |
109 101 |
39 62 |
25 17 |
36 21 |
564 528 |
26 1 |
84 63 |
12.0 12.5 |
66 68 |
| 75 | 有 無 |
100 100 |
37 44 |
15 29 |
48 27 |
627 581 |
35 24 |
88 87 |
11.2 11.7 |
80 81 |
| 100 | 有 無 |
111 101 |
32 47 |
14 18 |
53 35 |
522 582 |
27 7 |
91 90 |
11.4 11.7 |
80 83 |
春になれば、自然温度にまかせたままの越年蚕種でも、それぞれの地方の桑の発芽とかなりよく歩調を合わせて胚が発育し、孵化するから(94,520)、自分の畑の桑と手持ちの蚕種とで蚕を飼うだけならば、自然放任で間に合う揚合もあるが、これでは、採種時期が異なり、産他の異なる(92)蚕種を予定の時期に掃立てることは不可能である。
活性化程度の異なる蚕種は、催青の段階でいくら手をつくしても、揃えて孵化させることはむずかしいから、活性化を揃えるために、産卵直談から、夏、秋、冬を通しての蚕種保護が大切で、この準備を抜きにして催青を考えることはできない。
蚕種保護の方法は採種時期によって違えなければならないが、これは貯蔵期間の長短、気温の高低なでに応じて、蚕種の休眠と健康とをどのように管理するかによるものであるから、貯蔵期間が最も長く、その間の気候の変化の最も激しい春採りの場合について委しく説明し、その他の時期に採種するものについては、その相違点だけを記述する。これまでに述べてきた基礎研究の成果をどのように実際の場面に適用するかがその主題である。
1 春採り蚕種
A 産卵から休眠の完成まで
蚕種保護と云えば収蛾後の問題のように考えられ勝ちであるが、実際には産卵中から始まる。高温、低温その他の外因に対して蚕種の抵抗力の最も弱いのは産卵後2時間目頃であるが(Ⅵ3Da)、この時期は母蛾に産卵させている間に経過するのが普通で、産卵中の不良環境のために死卵や不受精卵の生ずることがある(Ⅵ5Aa)。産卵中の保護温度は23℃を下らず25℃を超えない範囲に保つのがよい。これは蚕種保護のための必要を満たすと共に産卵能率を高め、ひいては人工孵化の成績を上げることにもなる。収蛾の頃の卵には所謂蚕卵抵抗力曲線の第2降下点前後のものが多いから、取扱いを丁寧にし、物理的な衝撃、例えば台紙の裏を強く叩いて蛾を払うようなことも避けなければならない。
再出卵の出るおそれのある場合には、産卵後12時間以内に15℃に移して5日間おけばこれを防止できると云うことが、殆ど常識的に云われているが、これはそのときどきの条件によって必らずしも効果が一定しない。また再出卵そのものの解釈も様々で、上記のような処置では全然防止できない再出卵のあることも考えなければならない。
産卵直後の15℃保護によって防止できるのは、親の代の越年性管理が不十分なために、越年性の不安定な蚕種ができ、着色卵として産み出されながら2週間以内ぐらいで孵化する特殊な再出卵(渡辺型)であるから(Ⅲ3Bc)、その正しい防止法は親の代の越年性管理を十分にすることで、産卵後の15℃保護は云わば補助手段に過ぎない。越年性管理を完全にして、このような処置の必要でない蚕種を作るのが本来の蚕種保護法である。
15℃で保護したものは、その後の高温保護に注意して、蚕種を軟性(Ⅴ2A)にしないように管理しなければならない。
産卵後の高温期間の不足なために生じた軟性卵は、産卵後1カ月以上もたってから不時発生を起こすことがある。これも再出卵と呼ばれている。この型(軟性卵型)の再出卵は、越年性の不十分な蚕種と云う点では渡辺型と同じであるが、親の代の越年性管理とは無関係に、産卵後の休眠完成の不完全な場合に、一般的に発生するもので、産卵直後の15℃
保護によっては防止できない。
なお、軟性卵と云われているものの中には、親の代の越年性管理の不十分が原因で、渡辺型の再出卵にまではならないが、生まれながら軟性卵になっているものが含まれている。このようなものに発生する再出卵は渡辺型の延長に過ぎないから、防止法は渡辺型の場合と同様である。
越年性卵が翌春の正常孵化以前に発蛾するものを、原因の如何に拘らず、総べて再出卵とする考え方もある。例えば、洗落しの刺戟による不時発生がこれである(1118)。
河合は十数篇に上る百出卵の研究を発表しているが(337-342,344,347,349-358)、その大部分は、洗落し後の越年蚕種を、胚の発育し得る温度で保護した場合の不時発蛾に関するもので、洗落しの刺戟による活性化の促進が原因になっている。このような再出卵は、産卵後を低温に保護しても(349)、産卵後25℃に35日間保護して硬性卵にしておいても(351)防止することはできない。
河合は、再出卵の発現を3・ヒドロキシキヌレニンの母体から卵内への移行、および卵内における漿膜色素形成のためのその消費の面から研究しているが、その結果からみても、産卵後2週間内外で孵化する種類の再出卵は、産卵直後に進行する越年性確定の生理作用が阻害されるために発現するものであるから、普通の品種に対しては、産卵直後の15℃、5日間の低温保護がその発現防止に有効である。
卵管各部の3・ヒドロキシキヌレニン量を測定すると、基部に多く先端部に少ない。再出卵は1蛾の卵のうちでもこの先端で作られたもの、換言すればおそく産み出されるものに多いと云われているが(636)、実際に産卵を調べてみると、必らずしもそうとは云われない成績も出ている(4,5)。
洗落しによる再出卵の研究からわかるように、物理的な刺戟によって越年性卵が活性化することが多いが、刺戟の弱い場合には再出卵にまではならずに、胚が臨界期頃まで発育して死んでいる場合も少なくない。これらは原因不明の死卵として方付けられがちであるが、原因は洗落しによる再出卵と同じであるから、保護中、特に越年性の不安定な産卵後10日以内は取扱いに注意し、振動、摩擦、異常な温度変化、その他総べての刺戟を避けることが大切である。これは卵ばかりでなく、蛹の時期についても同様である(Ⅴ1Ac)。
産卵直後からの多湿は高温および低温と相俟って蚕卵に生理的障害を与える。沓掛(第107表)および勝又(318)の成績からみて、産卵後、休眠完成までの保護温湿度は25℃、75%前後が適当と考えられる。
高島・神田(963)は、温度25℃で、産卵後40日までの間の保護湿度を比較し、40-90%の範囲内では翌年の孵化に大差はないが、どちらかと云えば90%がよかったと云い、足立ら(7,8)の成績によれば、7月下句から9月まで45日間の試験において、65-95%の範囲では85%(温度25℃内外)がよかった。
沓掛は孵化歩合ばかりでなく、孵化した蟻蚕の飼育試験も行ない、高温多湿の影響は稚蚕の減蚕歩合にも大きな影響をおよぼすが、壮蚕においては稚蚕におけるほど大きな影響がなく、繭質への影響は更に少なかったと云っている。ただ、沓掛の成績は高温30℃、低温20℃
と組合わせて多湿98%の影響を調べたもので、かなり極端な場合の影響を示すものである。 `
多湿には、卵の生理障害ばかりでなく、黴が生えたり、不受精卵や受精後間もない死卵が潰れずに残って、比重選や風選によって除き難い所謂赤死卵(ⅩⅠBa)を多くするなどの害もある。蚕種保護室を地下や半地下に設けると、温度の変化は少ないが、湿度の高いために赤死卵の多くなる気味がある。
勝又(318)の成績では湿度37%でも50%に劣ることのない孵化歩合を示しているが、漿膜または漿膜クチクラ完成前(Ⅵ2Ba)の卵が乾燥に弱いことは確かであるから、即浸前冷蔵の際などは注意を要する。直冷式の冷蔵庫は設計が悪いと非常に乾燥する。実験用に電気冷蔵庫を用いて蚕種を冷蔵する場合には、特にこの点の注意が大切である。
休眠完成のための最適温度は25℃、休眠完成に要する日数は約30日(Ⅴ1Ba)と考えられるが、原種においては、同時にこれが障害を来さない25℃日数の安全限界でもある。交雑種においてはこの安全限界は50-60日と考えられる。高温保護(25℃)日数はこの限度内において、蚕種の貯蔵予定期間に応じて調節するのであるが、翌年5月掃立て用蚕種ならば限度一杯の高温保護をする。貯蔵期間が長いから、十分に休眠させて、その期間を貯蔵に利用するのである。
春採り蚕種の夏秋期保護は自然温度で行なわれることが多いが、自然温度は同一地方においても年によってかなり相違し、それが蚕種の休眠完成、従って活性化の遅速に影響することが大きいから注意を要する。
第113表は新庄原蚕種製造所における7月から翌年1月前半までの蚕種庫内温度と、その蚕種庫内に保護した蚕種の1月15日の孵化歩合とを、3カ年に亘って調査したものである。
| 年度 | 7月 | 8月 | 9月 | 10月 | 11月 | 12月 | 1月 | |||||
| 後期 | 前期 | 後期 | 前期 | 後期 | 前期 | 後期 | 前期 | 後期 | 前期 | 後期 | 前期 | |
昭和34年 35年 36年 |
℃ 22.1 25.5 24.9 |
℃ 21.9 25.8 25.0 |
℃ 23.8 23.6 25.2 |
℃ 21.8 22.0 24.4 |
℃ 21.4 20.0 21.3 |
℃ 18.3 16.5 18.4 |
℃ 17.2 13.5 15.7 |
℃ 13.5 11.5 11.9 |
℃ 9.7 9.5 9.3 |
℃ 7.7 5.5 6.5 |
℃ 2.1 4.1 3.7 |
℃ 1.5 1.9 3.4 |
| 年 度 | 日122号 (%) |
日124号 (%) |
日125号 (%) |
瑞 光 (%) |
支25号 (%) |
支122号 (太)(%) |
支124号 (%) |
銀 白 (%) |
| 昭和34年 35年 36年 |
56 6 8 |
21 2 3 |
40 5 9 |
95 50 72 |
76 36 63 |
94 67 77 |
93 92 93 |
69 32 31 |
年による孵化の遅速(活性化の遅速)が夏期保護温度の高低だけによると云う根拠はないが、温度と孵化歩合とを対比すると、34年の夏期の低温が同年の活性化の旱かった原因の一つであろうと云うことが十分に考えられる。もし温度がこの原因であるとすると、34年の9月以降は35年および36年よりも温度の下り方が遅く、この時期の温度が活性化を促進したとは考えられない。また8月後期および9月前期の温度も35年と相違がなく、差の大きいのは7月後期から8月前期にかけての温度であるから、この時期の低温が休眠の完成に影響したものと考えられ、休眠の完成には産卵後1カ月間の温度管理の大切なことがうかがわれる。
最近、夏期の高温による蚕種の被害を防止するために、蚕種庫に自動調節式のクーラーが使用されるようになったが、この使用を誤まったためと考えられる蚕種の障害が注目され初めている。これは蚕保護上重要な問題であるが、冷蔵庫の管理とも共通する問題であるから、Ⅶ1Cにおいて述べる。
B 休眠完成から初冬期まで
春採り蚕種の休眠が完成するのは7月下旬乃至8月下旬で、最も暑い時期に当るから、そのまま自然温度におくと、25℃
以上の期間が加算されて孵化が悪くなる。高梨(954)は、6月上旬に採種した蚕種を9月20日まで25℃に保護する区と7月下旬から9月下旬にかけての平均温度が25℃を超える自然温度に保護する区とに分けて、翌春の孵化歩合を比較し、明かに後者の劣ることを示している。蚕種保護法の試験において、25℃、1カ月で自然温度に移したと云うような記載をみることがあるが、これでは25℃の恒温が1カ月であったと云うだけで、その蚕種が実際に接触した25℃以上の温度は、自然温度の分も加えると1カ月とは限らず2カ月であったかも知れない訳で、試験の条件としては極めて不明確である。
高温期間が長いと、品種によっては(支127号、銀白など)白ハゼ卵の多発することもある(1028)。 しかし、休眠完成後、直ちに次ぎ次ぎと保護温度を下げて行くと、活性化が早く、冬期の取扱いに困ることになるから、蚕卵の生理をそこなわず、また活性化を促進することもない23℃-20℃に保護し、蚕種庫の平均温度が20℃以下に下がるのを待って自然温度に移す(940)。20℃は夏期保護温度の中間温度と云うことができる。
10月初旬になっても平均温度が20℃以下に下らないような地方では、人工的に徐々に温度を下げて、12月中・下旬に5℃になるように管理するのがよい。
昭和32年度地方蚕業試験場協力試験として、越年種の秋期保護に関する試験が、全国17蚕業試験場の協力によって行なわれた。この試験の設計は第114表の通りで、試験結果から次ぎのように結論されている。
| 試験区 | 保護日数 | 冷蔵時期 および 方法 |
出庫時期 および 方法 |
|||
| 25℃ | 15℃ | 5℃ | 産卵から 冷蔵まで |
|||
| a区 | 産卵から 60日 | 25℃ 60日後 60日 |
15℃ 60日後 30-60日 |
150- 180日 |
低温に対する 抵抗力の最も 強い甲胚子に なった時に冷 蔵する。 |
各場所の春蚕掃立 期に孵化するように 出庫し、従来の各場 所慣行の方法で催青 し(各区同一方法)、 孵化調査をする。 |
| b区 | 同 90日 | 25℃ 90日後 60日 |
15℃ 60日後 30-60日 |
180- 210日 |
||
| c区 | 同 120日 | 25℃ 120日後 60日 |
15℃ 60日後 30-60日 |
210- 240日 |
||
| 対照区 | 産卵から12月末まで各場所の慣行保護温度で保護する | 180- 240日 |
||||
全孵化歩合および最多2日孵化歩合は、殆ど何れの場所においても対照区が比較的良好であるが、a区より劣る場合もある。a区はb区に勝り、一般にb区はc区に勝る傾向が認められるが、愛知と鹿児島とにおいてはこれと正反対の傾向を示した。長野、愛媛、熊本においては一定の傾向がなく、各区間に大差がなかった。
この結論は、結局、高温期間の長いのは悪いと云うことであろうが、各場所における実際の保護温度(設計通りには行なわれていないから)とその成績との関係の吟味、25℃から直ちに15℃に移して夏秋期を保護するような取扱いが活性化の早晩や冷蔵抵抗力にどのような影響をおよぼしたか、などの分析が行なわれていないために、試験結果を実際の上に生かすことのできないのは遺憾である。
11月から冬中にかけての保護湿度については、山口・清水の成績(第108表)においては90%の場合の孵化が最もよかったが、勝又(318)によれば26-90の範囲では90%が最も悪く、その他の区間には大差がないが、50-60%がよいように見受けられる。このように正反対ともみられるような成績の出ていることは、湿度以外の条件が大きく影響しているためで、湿度そのものの影響は比較的小さいことを示すものであろう。休眠完成までと同様、75%内外で差支えなく、特別に調節をする必要もないものと思われる。
11月下句から12月初句にかけて、バラ種別のものは洗落し(Ⅹ1A)、乾き次第、気温が5℃以上の地方では5℃に移す。洗落しの刺戟によって活性化の進んだ蚕種は、5℃以上の温度にあうと発育するおそれがある。
1蛾採りの蚕種は2%ホルムアルデヒド液に30分ぐらい浸漬して卵面消毒を行ない、水洗いし、乾かして5℃に移す。以前は寒中に種洗いをする習慣があったが、人工低温によって活性化を進める場合に、汚れたままの蚕種を冷蔵庫に持込むのは好ましくないから、洗落しと同じ時期に洗っておくのがよい。
C 人工温度について
蚕種保護室の温度調節は、室の構造や使用法からみても、飼育室に比べて遙かに容易である。殊に最近は新らしい調節装置ができて、この点の心配は殆どないように考えられがちであるが、実際には必らずしもそうではなく、新らしい装置を採用したための問題も起こっている。工学的な問題はその方面の専門家にまかせるとしても、装置に対する希望条件を出し、選択し、使用して行く上での蚕種技術者の責任は、従来に増して重くなっていることを考えなければならない。
例えば、風穴を利用していた頃には、自然温度をそのまま利用するのであるから、適当な場所を選ぶことに問題はあるが、場所がきまってしまえば、それを用いて行ない得る蚕種保護には限度があって、自然に頼ることが多く、蚕種技術者の責任範囲は比較的狭かった。氷庫を使う場合には、氷で設定することのできる温度の範囲はきまっていた。
アンモニア式冷蔵庫を使用するようになると、保護温度をどのように設定するかは完全に蚕種技術者の責任になり、温度調節の可能範囲が拡まっただけに、温度の選択、組合わせを技術者の責任において決定しなければならなくなった。
フレオンによる自動調節では、機械の操作は確かに容易になったが、アンモニア式の場合よりも遙かに低温の空気が室内を流れるから、平均は5℃であっても、冷蔵庫の構造や冷却パイプの配置などが適切でないと、目的温度以下のつめたい空気が頻繁に触れるようなことも起こる。
気温30℃のときに室温を25℃に保つために、25℃の空気を作って室内に送り込めば問題はないが、それでは経費がかさむので、普通には非常につめたい空気で冷やしている。人間の場合には衣服を調節するとか 居場所を変えるとかして不快さを避けることができるが、それでも体の調子をそこねることがある。飼育室の場合には、1日に何回か給桑のために出入りするから、寒暖計ばかりではなく、食桑状態、蚕の動きなどの生物的な反応を通しても、環境の適否に早く気付くことができるが、蚕種はこのような反応をそのときどきに示すことがないために、寒暖計だけが頼りである。
長い保護期間に亘って、特定の場所に設置した寒暖計によって機械的に観測を続けているうちには、細かい注意のなおざりになることもあろうが、それよりも、設定した温度の蚕種に対する適否、および寒暖計が蚕種の接触する温度を正しく示すように配置されているか否かが問題で、装置そのものよりも蚕種技術者の責任に属する問題が多い。
自然温度の平均25℃と28℃との場合には、何れも上下に巾のある温度で、かなりの部分で互に重なり合っているから、適温が何れの側にあるにしても、保護温度に対する蚕種の反応は連続的な変化として両区の成績に現われるが、恒温の25℃と28℃との間にはこの重なり合いが少ないか、あるいは全くないために、両区の成績の違いがはっきり出る。平均気温はかなり違っていても、自然温度での蚕種の活性化時期に毎年極端な違いのないのは、この重なり合いのあるためと考えられる。これに対し、人工温度によって比較すると、2℃-3℃の温度差も明瞭な結果の違いになって現われることが多い。正確な温度調節ができるようになればなるほど、ごまかしがきかなくなって、技術者の責任が重くなる。
活性化について云えば、5℃でも7℃でも、あるいは3℃でもあまり遅速なく活性化する品種もあれば、5℃より低いと目立って活性化のおくれる品種もある。支124号は前者、日124号は後者の例であるが、このような違いは珍らしいことではない(685,742,769,936)。日129号および支129号の温度別活性化速度を第115表に示す。
| 蚕品種 | 冷蔵日数 (日) |
催青2週間孵化歩合(%) | |
| 5℃ | 7.5℃ | ||
| 日129号 | 40 60 90 |
0 0 30 |
0 2 75 |
| 支129号 | 40 60 90 |
0 1 53 |
0 11 91 |
| 日129号×支129号 | 40 60 90 |
0 0 80 |
0 2 87 |
| 支129号×日129号 | 40 60 90 |
0 0 39 |
0 6 92 |
冷蔵車内の上部と下部とで2℃-3℃の温度差のあることは珍らしくないが、同一品種でも、上段のものと下段のものとで活性化に遅速の生じ得ることはこの表によって明らかであろう。
催青室で蚕種の積みかえを行なうように、毎日の必要はないが、室内温度の分布によっては、冷蔵庫内の蚕種もときどき積みかえを行なえば、活性化を揃えるのに有効である。
人工的に温度を調節する場合の蚕種庫や冷蔵車内の温度分布は、外温の高低によっても、室内に物を入れた場合と空の場合とでも相違するから、棚の配置、蚕種の収容量などを予め考慮して設計しておかないと、装置が出来てから不備に気付くことがある。
蚕種庫の冷房を誤まったための蚕種の障害には、温度分布についての不注意、休眠完成後の温度の下げ方が早過ぎたこと、目的温度のきめ方を誤まったこと、などが原因になっている場合が多い。
人工温度を設定する場合の基礎資料として、恒温と変温(平均は恒温と同じで上下に変動のある温度)との蚕種におよぼす影響を明かにしてお<必要がある。例えば、夏期高温の地方においての、休眠完成後の人工温度を東北あるいは長野の温度にならって設定しようと考えた場合に、これらの地方の月平均あるいは旬平均をそのま恒温に直して適用しても、果してこれらの地方における成績と同じ結果が期待できるかどうかと云うことである。完全な恒温を作ることは実用上できないから、恒温と云い変温と云っても程度の問題に過ぎないが、正確な温度調節ができるようになるほど、重要な問題になってくる。現在、この点についての信頼のできる成績は殆どない(200,235,236)。蚕種保護に限らず、平均温度によって簡単に結果が比較されがちであるが、平均によって比較してもよいのはどの程度の変温までかと云うことが明かでない。催青温度についての恒亘温と変温とを比較した平山の成績(Ⅸ2A)なども、この点から一層の検討が望ましい。恒温と変温とに対する蚕種の反応は、休眠完成、活性化、催青など、それぞれの場合によって異なるのではないかと考えられる。
一般に、建設関係者は、冷蔵庫と云えば魚類や野菜の冷蔵庫、冷暖房と云えば自己防衛の可能な人間の場合の経験によって設計しがちで、これに予算の問題も関係して、蚕種技術者の希望の容れられない場合が多いが、正しい要求の主旨を理解させてゆくことも蚕種技術者の責任である。フレオンによる直冷式に改めたために、冷蔵庫が乾燥して使用できなかったと云うような実例は、建設費の節約によって湿度に対する希望の無視された例と云えよう。
D 越冬期
越冬期の保護法は 単式冷蔵、中間手入れ式冷蔵、および複式冷蔵の三つに大別することができる。中間手入れ式は複式冷蔵と原理が同じなので複式冷蔵と呼ばれることも多いが(第114表)、元来の複式冷蔵とは取扱いも目的もかなり相違しているからこれを同じ名前で呼ぶのはまぎらわしい。
a 単式冷蔵法
本来の単式冷蔵は、越年蚕種を蚕種庫内の自然温度で保護し、胚が丙A-丙Bに発育したとき2.5℃に冷蔵し、催青着手までおく方法で、寒地においては普通に行なわれている。胚が丙Bを過ぎると冷蔵に耐える力が低下するから(第119表)、丙Bを過ぎないように、丙A-丙Bで冷蔵する。東京付近の気候では、例年、2月下旬から3月上旬にこの時期になる。洗落し後も5℃以上の気温が続くような場合には、5℃の冷蔵車に移して胚の不時発育を防止する。不時発育を始めた卵は春までの間に死卵になることが多い。
暖地においては、蚕種の活性化がおそいものと考えられがちであるが、一部の卵は案外早く不完全な活性状態になり(335)、蚕種全体としての活性化が不揃いである。早く活性化したものは気温が高いと徐々に発育を始めるが、春の催青までに活力が低下して孵化不良の原因になる。暖地の蚕種の孵化が悪いのは、夏秋期の高温の害と共に活性化の不揃いが主な原因であるから、このような地方においては、夏秋期の温度管理を行なうと共に、洗落し後は直ちに蚕種を5℃に保護し、活性化の揃ったときに(中間手入れ式の項参照)2.5℃に冷蔵する。
渡辺(1151)は、伊豆の松崎の自然温度で保護中の蚕種の一部を種々な時期に東京へ送って、東京の自然温度で保護したものと松崎に残したものとの4月になっての孵化歩合を比較した。それによると、総孵化歩合には差がなかったが、最多2日孵化歩合には、1月1日以前に東京へ送ったものは高く、その後急に低下することが認められた(第116表)。東京から松崎へ送って松崎の自然温度で保護した卵は、これと反対に、1月1日以前に送ったものの最多2日孵化歩合が劣り、以後急に高まった。
| 蚕品種 | 発送月日 (月・日) |
松崎から東京へ送った卵 | 松崎に残した卵 | ||
| 総孵化歩合 (%) |
最多2日孵化 歩合(%) |
総孵化歩合 (%) |
最多2日孵化 歩合(%) |
||
| 支108号 (6月9日産卵) |
9・5 10・1 11・1 12・1 1・1 2・1 3・1 4・1 |
91 91 89 89 90 91 93 92 |
90 89 87 84 63 73 72 71 |
91 90 85 85 90 91 90 89 |
46 60 45 57 57 56 52 63 |
| 支108号 (8月29日産卵) |
9・1 10・1 11・1 12・1 1・1 2・1 3・1 4・1 |
97 98 98 97 97 94 97 97 |
93 98 97 96 71 75 78 62 |
93 96 96 96 97 95 95 95 |
65 68 56 46 57 54 60 57 |
渡辺は、松綺で保護して1月以降に東京へ送ったものの孵化が不斉になるのは、暖い松崎に1月以後まで保護すると、活性化に必要な5℃以下の温度に接触する日数が不足するためであると考え、更に、全国各地の蚕業試験場における、冬期間5℃以下の温度に接触する日数と越年蚕種の孵比の斉否との関係を調査した結果、1月以後に平均5℃以下の温度に接触する日数が、大約24.5日以内の地方においては孵化が甚しく不斉であり、25-50日の地方においては孵化の斉一な場合と不斉な場合とがあり、50日以上の地方においては孵化の斉一な場合の多いのが普通であると結論した。
河合(336)も新潟から東京へ蚕種を送って同様な試験を行ない、渡辺と同じ結果を得たと報告しているが、河合の成績の中には、渡辺の説明だけによっては理解し難い問題が含まれているように思われる。
渡辺の成績においては(第116表)、最多2日孵化歩合の悪くなる境目と云われる1月1日送付の分の最多2日孵化歩合が、その前後の分に比べて特に落ちているが、この低下は総孵化歩合にはみられず、また東京から松崎へ送ったものにも認められなかった。処が、河合の成績においてはこの現象が極めて明瞭で、総孵化歩合にも、新潟に残した分にもはっきり現われている(第117表)。河合はこのほかに支115号の夏採り蚕種の送付をも行なっているが、これにも同様な現象が認められる。
| 発送月日 (月・日) |
東京へ送った卵 | 新潟に残した卵 | ||
| 総孵化歩合 (%) |
最多2日孵化 歩合(%) |
総孵化歩合 (%) |
最多2日孵化 歩合(%) |
|
| 9・ 1 9・25 10・25 11・25 12・25 1・10 1・25 2・10 2・25 3・10 3・25 4・10 4・25 |
64 85 78 82 64 51 69 79 63 86 84 92 96 |
62 72 61 67 63 49 65 73 55 83 70 79 91 |
95 93 96 96 98 91 90 88 96 98 91 97 93 |
93 93 95 96 95 89 90 87 93 95 90 96 93 |
松崎の場合だけならは偶然と考見られたのてあるが、活性化の十分でなかった筈の10月、11月よりも、寒い新潟においては活性化の進んだ筈の12月、1月、2月の蚕種を送った成績の方が悪かったと云う河合の結果を考え合わせると、単なる偶然として方付けることはできない。しかも新潟に残したものにも同じ影響が現われているのである。
前もって蚕種庫内で切分けておいた台紙片を、送付のたびに取出したのか、送るたびに蚕種を実験室に持ち出して切分けたのかは記載されていないが、もし後者であるとすると、蚕種庫からの出し入れの間に、送った分と残した分とに共通なある原因が働き、送った分には、その影響を更に強めるような条件が加わったものと考えなければならない。
推測の範囲を出ないが、この点に最も関係が深いと考えられるのは、冬期間に蚕種を不時高温に接触させた場合の障害である(419)。越年蚕種に対する不時高温の害の最も大きいのは越冬期であることを考えると、上記の成績はこの点から再検討を加える必要がある。松崎と違って新潟は冬期の気温が低く、実験室にも郵便局にも、また輸送中の車内にも暖房のあることが考えられるから、蚕種庫からの出し入れによる温度較差に郵送中の暖房の影響が加重すれば、東京へ送ったものの障害は新潟に残したものにおけるよりも大きい筈である。
河合は同じ論文の中で、自然温度で保護中の蚕種を、種々な時期に10日間25℃に接触させた後、再び自然温度に戻しておくと、春になっての孵化歩合は、12月25日から3月25日頃までの間に高温にあわせた区が悪かったが、5日間の接触では影響がなかったと云う実験結果を述べている。活性卵を5日、10日と云うような長い期間高温にあわせると発育が進むから、これを低い自然温度に戻すと、胚の発育段階別の冷蔵抵抗力にも関係し、不時高温とは別の問題が加わってくる。谷口(1038)は11月20日から12月20日まで1カ月間の25℃接触を試験している。
普通に冬期間の不時高温の害と云うのは、高温接触1日内外の短時間の場合か多く、接触期間が長いと却って被害の軽減するらしい傾回さえみられる。その原因は明かでないが、臨界期になると不時高温の被害が減少するから、休眠が終って発育の始まろうとする前には、冷蔵浸酸種の再冷蔵による白ハゼ卵の発現時期にもみられるように、何か生理的に不安定な状態が存在するのであろうと思われる。
寒地の自然温度においては、蚕種は揃って早く活性化すると云われてはいるが、品種により、採種時期によって(高温期間が異なる)、同一品種の間にもかなりの活性化の遅速がある(第118表)。
| 採種 時期 |
蚕品種 | 11月15日 | 11月30日 | 12月15日 | |||
| 孵化歩合 (%) |
孵化期間 (日) |
孵化歩合 (%) |
孵化期間 (日) |
孵化歩合 (%) |
孵化期間 (日) |
||
| 春 | 日112号 日122号 支110号 支108号(旧) |
0 0 27 17 |
- - 14 17 |
64 5 79 77 |
18 6 22 20 |
75 49 83 81 |
17 14 15 15 |
| 秋 | 日112号 日122号 支110号 支108号(旧) |
0 0 0 2 |
- - - 4 |
5 3 93 89 |
5 9 16 17 |
47 47 82 93 |
15 14 15 16 |
寒地の蚕種は、活性化はしても、冬中寒さのために発育を抑えられている状態であって、形態的にはおそくまで丙Bにならないが、早く活性化しているため、長い越冬中に冷蔵に耐える生理的限度を超えてしまい、孵化歩合の低下していることがある。このような胚は春になって気温が上ると忽ち丙Bになるが、このときになって冷蔵してみても、一旦低下した孵化歩合はもとには戻らない。先きに(Ⅵ2Bd)胚の形態と生理状態とのずれとして述べたのはこのことであって、胚の形態ばかりを頼りにすることはできない。
この例は第113表に示した支25号にみられる。この品種は冷蔵に耐える期間が短かいように考えられるが、特に夏期の保護濃度の低かった昭和34年には、冬期の蚕種庫内温度が、1月前期15℃、後期-0.4℃、
2月前期1.2℃、後期2.0℃、3月前期3℃、後期3.9℃、4月前期5.3℃
と、かなり低かったにも拘らす、孵化歩合が最高を示したのは3月15日出庫の99%で、以後低下し、4月30日の出庫では45%に落ちていた。しかも胚の形態は+乙Bに留まっていた。寒地の蚕種保護には心配がないと云われているが、冬の長い地方においては、これに似た傾向が他の品種にも認められることが少なくない。このような傾向のある地方では、夏秋期の濃度管理によって活性化をおくらせるとか、活性化したら直ぐ0℃に冷蔵するとか、中間手入れ式または複式冷蔵法を採用するとかの対策を講ずる必要がある。
単式で蚕種を長期間冷蔵するために、1月頃から0℃に近い温度に冷蔵する方法が以前は行なわれた。秋採りの支101号をこの方法によって翌年の秋蚕用に冷蔵すると、他の品種に比べて孵化歩合の悪いことが、大正9年頃に問題になったが、調査の結果、これは支101号の冷蔵抵抗力が弱いためではなく、この品種の胚が早く発育を始めるため、他の品種と同じ扱いをすると、冷蔵時期のおそ過ぎることになるのが原因であると報告されている。渡辺(1138)によれば、1月25日には共に休眠期(甲胚子の意味)であったが、2月25日には、支101号においては最長期末期まで発育しており、日106号においては、休眠期よりは幾分長いが、最長期には達しない状態に止まっていた。これらの卵を33゚F(0.6℃)に冷蔵し、7月10日に出庫、催青した処、孵化歩合は5蛾平均において、1月25日冷蔵のものは支101号92.7%、日106号95.8%、2月25日冷蔵のものは支101号81.0%、日106号95.3%であった。
1月から40゚F(4.4℃)に保護してそのまま4月までおく方法、1月は40゚F(4.4℃)2月は38゚F(3.3℃)、3月40゚F(4.4℃)、4月45゚F(7.2℃)と変化させる方法なでも行なわれたことがあるが現在では用いられていない。
鈴木(893)の測定した冬期間の蚕種の炭酸ガス排出量を用い、先きに述べた方法(Ⅵ3Ba)によって、冬期間の越年種の所要空気量を、室賀は蚕種1kg当り15.19㍑(5℃の場合)と計算した。しかし、荒木ら(49)が、容積1㍑のブリキ罐に350蛾分の蚕種を容れ、パラフィンで密封したものは、湿度が37゚F(2.8℃)以下の場合には2月から8月まで貯蔵しても害がなかったと云う結果から1蛾の卵量を0.3gとして計算すると、所要空気量は蚕種1kgにつき全期間9.9㍑で足りることになり、東畑が、冷蔵は1㍑に蚕種4.5g以下としているのから計算すると、冷蔵期間中を通じて蚕種1kg当り22.2㍑の所要空気量になると云う。計算値は、不越年種について述べたように、十分に余裕のある量と考えられる。冷蔵中の蚕種貯蔵罐は銹びていても害がない。
b 中間手入れ式冷蔵法
冷蔵に耐える期間の限度に近付いた蚕種を冷蔵庫から出して、胚の発育をある程度進めると、その蚕種は再び冷蔵に耐えるようになる(Ⅶ1Dc)。この性質を利用し、蚕種の貯蔵期間を延長すると共に、その後の取扱いを便利にするために、冷蔵期間の途中で蚕種を一度出庫し、胚を丙A-丙Bまで発育させた後、再び冷蔵するのが中間手入れ式冷蔵法てある。例えば、第28図の例において、2.5℃区の卵に4月6日から11日まで10-15℃で中間手入れを施し、90%の胚を丙A-丙Bに進めた後、2.5℃に再冷蔵したものと中間手入れを施さずに2.5℃に冷蔵し続けたものとの9月1日出庫の孵化歩合は、初発3日孵化歩合が2.5℃連続区71%、手入れ区87%、催青2週間孵化歩合が前者84%、後者90%であった。
中間手入れ式冷蔵は 1)低温保護、2)冷蔵、3)中間子入れ、4)再冷蔵の4段階の処理によるものである(第29図)。

第29図 中間手入れ式単式冷蔵法模式
a:活性化を進めるための低温保護(5℃)、b:活性化した蚕種の冷蔵(2.5℃または0℃)
c:中間手入れ(10-15℃)、d:再冷蔵(2.5℃)
第1段階の低温保護は蚕種の洗落し直後から始める。洗落さない蚕種についても同様に12月初、中旬から始める。温度は5℃である。
中間手入れ式冷蔵の成績は、手入れを行なうときに蚕種が揃って完全に活性化しているか否かによってきまる。中間手入れを施した蚕種は孵化が揃うと云われているが、活性化が揃っていなければ中間手入れを行なうことができない。洗落した卵を直ちに5℃に保護するのは、洗落しの刺戟によって活性化した卵の不時発育を防ぐ意味もあるが、同時に、活性化の進んだものの発育を抑え、その間におくれたものの活性化を促進して、蚕種全体としての胚発育の歩調を揃えることに重要な意味がある。洗落さない蚕種についても5℃保護の意味は同様であって、中間手入れ式冷蔵における蚕種の低温保護は、単なる冷蔵ではない。この目的の低温保護には5℃が最も適している(Ⅴ1Bb)。低温保護は、活性化させるのが目的であって、胚を発育させるためではない。胚が発育すると中間手入れの効果は減少する(Ⅶ1Dc)。寒地においては中間手入れの効果がないと云われることがある。これは、春蚕用のように冷蔵期間の余り長くない場合には、単式冷蔵によっても、普通の品種はよく孵化するので、差の明かでないことも一つの原因であるが、低温保護中に胚の発育の進むことも原因になっているものと考えられる。
寒地において、第29図に示した公式をそのまま当てはめ、自然温度において既に活性化の初まった蚕種を12月初、中旬から5℃に移して1月下旬までおくと、胚の発育はかなり進む。これを更に3月下旬まで2.5℃に冷蔵した後、中間手入れを行なっても、単式冷蔵に比べて特に効果を期待することはできない。
低温保護中に最も大切なのは、活性化が揃い次第第2段階の冷蔵に移して、この間に胚の発育を進めないように注意することである。低温保護の期間は、普通の場合、洗落さない蚕種に対しては60日、洗落した蚕種に対してはその刺戟による活性化促進を考慮して40日が限度で、これ以上5℃におくと、活性化の進んだ胚が発育し、結果が不良になると云われている。しかし、これは完全活性卵になっていない蚕種を冷蔵した場合のことて、未発育ではあっても完全に活性化したもの(甲)は、5℃では間もなく発育を始める(第102表)。
活性化状態は孵化調査によって調べれば最も確実ではあるが、これには日数がかかり、その間にも活性化は進むから、調査の結果と現在の活性化状態との間にずれが起こる。
卵を解剖して胚を調べても、発育を始めていない胚の活性化状態を形態によって判断することはむずかしいから、数日間催青して、活性化した胚を発育させた上で調べる方法が行なわれている。
古和田(436,442)は、解剖する前に、蚕種を17.5℃に5日間保護した後に胚を調べ、保護中に、1日に水野の1階程ずつ発育が進んだものとみなし、5階程さかのぼって判定する方法を提唱している。また、比重1.10の塩酸で110゚F(43.3℃)、4分間の浸酸を施した後、17.5℃に5日間おいて調べると、発育するものとしないものとの開きが一層大きくなって、活性化の違いが正確に鑑別てきるとも云っている。
沓掛・黒岩(479)は、15℃に3日間保護した卵を剖検し、水野の丁A以上に発育した胚の割合(%)を求め、丁A発育度と名付けた。この値はその蚕種の最多2日孵化歩合と強い相関があり、丁A発育度45を示す時期が蚕種を5℃から2.5℃に移す適期であるとしている。 12月中旬に洗落して5℃に保護した春採り蚕種は2月上旬、同じく初秋および中秋採りは2月中旬、晩秋採りは3月上旬にこの時期になるのが普通で、丁A発育度45のときの孵化歩合は70%内外であると云う。但し、春月×宝鐘およびその反交についての調査である。
採種時期による活性化の遅速は高温保護日数によって相違し、必ずしも上記の順序とは限らない。
検知管を用いて蚕種の炭酸ガス排出量の簡易測定を行ない、その値によって活性化の程度を知ろうとする試みもあるが(872,943)、この場合にも出庫直後では殆ど信頼のできる結果は得られない。出庫後25℃に1日おくと、測定値と胚の発育との間にかなり高い相関が認められる。出庫後25℃、24時間の蚕種1gを100ccの容器内に1時間(25℃)収容したときに、容器内炭酸ガス濃度の増加が300-500ppmを示す場合には、その蚕種の催青3週間孵化歩合は80%以上であった。しかし、これは上記丁A発育度に比べて、活性化の測定としては一層間接的な方法である。
その他、活性化に伴なう卵汁水素イオン濃度の変化(148,625,965)、胚細胞の核仁の変化(669,1066,1068)、卵黄細胞の核濃縮(972)などの現象の応用も考えられるが、実用には結び付いていない。
結局、実用的な低温保護法としては、洗落した蚕種を乾き次第5℃に移し、40日(洗落さないものは60日)を限度に冷蔵するのであって、この間に胚を調べて、活性化の早いものは早く冷蔵する。
冷蔵のための温度は、完全に活性化した蚕種ならば、2.5℃よりも0℃の方がよい。これは、2.5℃よりも0℃の方が活性化した胚の発育を抑える(第102表)と共に有効冷蔵期間も長い(第119表)からである。
| 胚の発生段階 | 冷蔵温度 | |||
| -2.5℃ | 0℃ | +2.5℃ | +5℃ | |
甲 乙 A 乙 B 丙 A 丙 B 丁 A 丁 B |
日 170 120 70 70 60 60 30 |
日 150 120 70 100 60 60 30 |
日 150 70 70 90 100 80 30 |
日 120 70 70 40 30 30 30 |
しかし、低温保護中に総べての卵が揃って活性化しているとは限らないから、2.5℃に冷蔵して、冷蔵と共におくれたものの活性化を補なうのが普通である。
2.5℃冷蔵の限度は60日で、それ以上の冷蔵を必要とする場合には0℃に移す。
中間手入れは冷蔵しておいた蚕種を出庫して、胚を丙A-丙Bの段階まで進める燥作である。丙A-丙Bになった胚は冷蔵に耐える期間が長いから(第119表)、その範囲内では、中間手入れは何時行なってもよさそうに思われるが、実際の成績では催青着手の1カ月前ぐらいに行なった場合の孵化が最もよい。従って、5月の掃立てのためには、3月中・下旬に行なうのが普通である。
胚を丙A-丙Bまで進めるのは次ぎのような理由によるものである。
1)丙Bになると、はっきりした形態的特徴が現われて、胚の発育段階を見誤まることが少ない。
2)胚を丙Bまで進めておくと、その後、孵化までの催青所要日数が10日余りで、品種による差も少ない(第154表)から、催青計画を樹てるのに都合がよい。
3)冷蔵に耐える期間が長い(第119表)。
丙Bに揃えようとすると丙Bを過ぎるものもできるから、丙Bを過ぎないように丙A-丙Bを目標に手入れをする。手入れの温度は10-15℃がよい。手入れをこのような低温で行なうのは、若い発育段階の胚は、進んだ段階のものに比べて、低温における発育が比較的速い(第103表)性質を利用して、胚の発育を揃えるのが主要な理由であるが、冷蔵温度との較差が少なく、生理的に安全である。また手入れの実施面からみても、低温の方が胚の発育がおそく、丙Bを超えさせる危険が少ない。甲から丙Bまでは15℃で4、5日、10℃ては10日ぐらいかかるのが普通であって、10℃以下での手入れは日数がかかり過ぎて実用にならない。
手入れ中は胚の進み具合を毎日調べて(Ⅶ3A)、丙A-丙Bで2.5℃に再冷蔵する。丙A-丙B期の蚕種は2.5℃において有効冷蔵日数が最も長い(第119表)。再冷蔵中の温度が2.5℃以上に、ならないように注意する。
c 複式冷蔵法
蚕種を、産卵後1年以上貯蔵する目的に使われる冷蔵法である。長い高温期間を経過している春採りよりも、初秋採りの方が冷蔵に耐える。
胚が未だ発育を始めない甲胚子の時期(普通1月中)に-2.5℃に冷蔵し、5月下旬に、一旦出庫して17.5℃に3-4日間おき、胚を丙A-丙Bに進めた後0℃に再冷蔵する(575,580)。
これは、甲および丙A-丙Bの有効冷蔵日数と最も有効な冷蔵温度とを組合わせて長期冷蔵を図るものであるから、種々な組合わせが考えられるが、上に挙けたのは最も成績のよかった組合わせの一つである。
具体的に示すと水野(580)は、8月採りの二化性中巣に、翌年1月20日に甲胚子の状態での第一次冷蔵(-2.5℃)を行ない、中間手入れを施こして、5月20日に丙Aで第二次冷蔵(0℃)を行なった。結果は、次ぎの通りであった。
7月20日出庫 孵化歩合 94.4%(0.54日)
8月20日 〃 〃 92.4%(0.43日)
9月20日 〃 〃 93.0%(0.63日)
括弧内は孵化日数の平均偏差
第二次冷蔵の温度を+2.5℃にしても成績はこれに劣らない。
これに対して、同じときから第一次冷蔵を行なったが、ただ手入れの時期を1カ月おくらせて、6月20日に第二次冷蔵を行なったものは
7月20日出庫 孵化歩合 62.1%(0.72日)
8月20日 〃 〃 89.5%(0.55日)
9月20日 〃 〃 90.1%(0.70日)
で、5月20日のものに比べて成績が悪かった。これは、手入れまでの第一次冷蔵(-2.5℃)期間が有効冷蔵日数の限度(第119表)に近付いたためで、限度を超えてから中間手入れを行ない、7月20日に第二次冷蔵を行なったものの孵化歩合は更に悪く、9月20日の出庫において79%であった。
第一次冷蔵の温度を0℃またはそれ以上にすると、冷蔵温度の組合わせ効果が少なく、長期の冷蔵に耐えない。これは冷蔵温度が高いと冷蔵中に甲胚子が乙または丙Aなどに発育するためで、例えば乙に発育した胚に手入れを施して丙Aまたは丙Bに進めても、手入れ前後の胚の形態に大差がないために効果がないのであろうと説明されている。胚の形に大差がなけれは何故効果がないのか、釈然としない説明てはあるが、手入れの効果の挙がらないのは事実である。
複式冷蔵法を利用し、2回以上手入れを繰返えして、蚕種を2カ年間貯蔵しようとする試験が行なわれたが、孵化歩合が著しく低く、実用にはならない(93,285,826)。これは、複式冷蔵の原理は、胚の発育段階別の耐冷蔵期間の組合わせであると云われていることから考えても当然の結果で、手入れを繰返えして胚の発育階程を進めれば進めるほど、有効冷蔵期間は短かくなるおそれさえある(第119表)。
2 初、晩秋および初冬採り蚕種
A 初、晩秋採り蚕種
8月下旬産卵の越年蚕種を翌年5月上旬に掃立てるためには、高温保護期間を春採りよりも短かくし、25℃期間を原種、交雑種ともに30日とする。その後は蚕種庫の温度によって、20℃を通し、または通さずに、自然温度に移す。以後の取扱いは春採りに準ずる。
10月中旬採りの越年種を翌年5月上旬に掃立てる場合には、25℃期間を20-25日とし、その後は1週間に5℃ぐらいずつ温度を下げて自然温度に移す。以後の取扱いは春採りに準ずる。
晩秋採りの蚕種は5℃に冷蔵して後、活性化の揃ったときに(春採り、中間手入れ式の項参照)2.5Cに冷蔵するだけで、中間手入れ式によらない場合が多いが、これは晩秋採りの活性化が春採りよりもおそく、2.5℃に冷蔵する時期が3月になるような場合の取扱いである。
仲野・北田(661)は、11月下旬採りの蚕種について試験を行ない、4月下旬の孵化成績は、高温(75゚F=23.9℃)期間20日、中間温度(60゚F=15.6℃)2日で自然温度に移し、3月中旬から41゚F(5℃)に冷蔵したものが、高温30日および10日のもの、冷蔵温度36゚F(2.2℃)および45゚F(7.2℃)のものに比べて最もよかったと報告している。晩秋採りても、冬の長い地方では、高温期間が足りないと、冬中に孵化歩合の低下することがある。要は、活性化と有効冷蔵期間との兼ね合いである。
古和田(441)は、10月採り蚕種の高温(25℃)保護終了後を、春採りのその時期以後と同じに取扱った場合、3月22日の出庫において、催青日数および催青積算温度(5℃以上)が春採りと全く同じになったのは高温30日の蚕種であったと云っているが、これは春採りの保護条件によっても相違する筈である。
これらの例によってわかるように、晩秋採りの高温期間とは云っても、その後の取扱いおよび催青までの冷蔵期間によって結果が相違する。小池・飛山(412)は、10月採り蚕種の高温期間を、翌年4月以前に掃立てるためには10日、5月中の掃立てには15-20日、6月以後の掃立てには20-30日にするのがよいと考えている。
梅原(1095)は、春蚕期に余った晩秋採り越年種を5℃に残しておき、初秋蚕に用いた処、孵化歩合は春採り即浸種に比べて3-6%ぐらい落ち、繭も軽目であったが、原種の保存、増殖には使用できると考えている。しかし、採種後どのような取扱いをした蚕種かを記載していない。
B 初冬採り蚕種
初冬採りの蚕種は増殖用の原種で、早春掃きに用いられることが多く、採種と掃立てとの間隔が60-150曰ぐらいの短期間であるため、普通の場合よりも産卵後の高温保護期間を長くした、特殊な冷蔵浸酸種として保護される場合が多い。特殊な用途であるため定法がなく、各自それぞれの見解によって行っている現状である。
初冬採りの卵は小形で、浸酸刺戟に弱いと考えられるが、このような卵を長期間冷蔵した後浸酸してよく孵化させるためには、高温期間を何日にするのがよいかが主要な問題点になっている。
緑川・近藤(528)は12月中旬採りの蚕種を用いて試験を行なったが、その結果から適当と考えられた高温保護期間は第120表の通りである。
| 出庫月日 | 冷蔵日数 | 蚕品種 | |
| 支那種 | 日本種 | ||
3月10日 3月30日 4月20日 |
5℃ 日 + 2.5℃ 日 日 30 + 40 = 70 30 + 60 = 90 30 + 80 =110 |
日 5 5-7 7-13 |
日 5-7 5-9 7-15 |
この場合は、所定日数の高温保護(25℃)の後、20℃、15℃、10℃各2日を経て5℃に移し、5℃30日で2.5℃に冷蔵したのであるが、別に行なった25℃で3-9日間保護した後、15℃1日を経て5℃に移し、5℃100日で2.5℃に冷蔵した試験成績は、これに比べて孵化がやや劣った。塩酸は比重1.10、液温は46.1℃(第121表)が47.8℃よりもよかった。47.8℃では刺戟が強すぎる。また出庫後、直ちに浸酸するよりも10℃、15℃各1日の中間温度を経て浸酸した方が成績がよかった。
| 出庫月日 | 冷蔵日数 | 蚕品種 | |
| 支那種 | 日本種 | ||
3月1日 4月20日 |
5℃ 日 + 2.5℃ 日 日 30 + 33 = 63 30 + 84 =114 |
分 4-5 3 |
分 5-6 3-4 |
しかし、初冬採り蚕種の性質はそれぞれの場合によって相違するため上とは違う保護法も行なわれている。古和田(440)は12月10日採種の春玉を、産卵後24-25℃に3日間保護した後5℃に64日間おき、2月23日に出庫し、比重1.10の塩酸で45℃、12分間の浸酸を行ない、浸酸後15.5℃、2日間の中間温度を経て25℃で催青したが総孵化歩合97.9%、最多2日孵化歩合92.7%であった。
小池・飛山(412)によれば、12月採り蚕種に対する地引氏の保護法は次ぎの通りである。
翌年4、5月に掃立てる場合 産卵中から24-25℃(湿度80%)で保護し、産卵後満7-10日間をこの温度においた後、20、15、10℃各2-3日間の中間温度を径て5℃に移し、翌春3月中、下旬頃に活性化するのを待って2.5℃に冷蔵する。掃立ての14-15日前に出庫し、冷蔵浸酸同様の浸酸を行ない、13-15℃の中間温度に4-5日間おいた後、催青する。
翌年2、3月に掃立てる場合 産卵中および産卵後を上と同じ温湿度で保護し、産卵後、2月掃きは満3昼夜、3月掃きは満4昼夜で5℃に移し、以後は冷蔵浸酸と同様に取扱う。
翌年6月以後に掃立てる場合 産卵中および産卵後の保護温湿度は上に同じ。産卵後この条件で10-20日間保護し、中間温度を経て5℃に移し、胚が丙Aになったとき2.5℃に冷蔵する。
初冬採り蚕種の保護法については、これまでに述べてきた越年性管理および後に述べる冷蔵浸酸の基本問題なとを考え合わせると、更に検討の余地があるように思われる。
3 蚕卵の簡易解剖法および孵化調査の注意
A 蚕卵の簡易解剖法
a 熱湯固定による方法
胚の解剖法には種々あるが、最も普通に行なわれるのは熱湯で卵を固定して行なう方法である。
1蛾採りの卵は台紙に着いたまま、バラ種はガーゼに包むか小さな金網の籠に入れて73-75℃の熱湯に3-5分間浸した後、水で冷やしてから乾かす。台紙が乾かないと、解剖のときに卵が台紙から離れ易く扱いにくい。バラ種は外面の水気を除いてから、スライドグラスの様なものの上にパラフィンを薄く塗った上にならべ、火の上にかざしてパラフィンをとかし、これに固着させる方法もあるが(723)、適当な堅さの紙片に糊を塗り、その上に揃べて乾かしてもよい。但し、固定した卵はあまり長時間乾かすと凹んてくるから、糊は乾き易いように濃いもの(ヤマト糊でよい)を薄く塗る。
台紙や糊が乾いて卵が固着着したならば、解剖針(木綿針を竹箸の先きに固着させたものでよい)の先きをよく研いだもの、または解剖刀(安全剃刀の刃を細かく折って竹箸に固着させたものでよい)を用い、反転前の卵ならば背側、反転後ならば腹側(第23図)の正中線を浅く切り破り、卵殼の片面を取除く。反転の前後で切り破る側を変えるのは胚を傷つけないためである(第24図)。
次ぎに、台紙または紙片を卵の付着したまま、シャーレまたは小皿に水を容れた中に沈め、スポイトで水を出し入れして煽ると、胚が卵殻および卵黄から離れて水中に出る。卵殻を切り破るとき漿膜が切れていないと胚が出ないから、最初の卵殻の切り方にも手加滅が必要である。深く切り過きると胚を損傷する。
このとき胚が卵黄に包まれて離れないのは固定濃度が高過ぎたか、時間の長過ぎ場合であり、胚が軟かく、千切れ易いのは固定の不十分な場合である。胚がよく固定されていて、卵黄のかたまらないのが適度の固定である。漿膜から1部分出ていながら胚の離れないときには、無理に水を煽ると千切れるから、針を使って離すのがよい。腹面に付着した卵黄は離れ易いが、背面の卵黄をスポイト操作だけで除くことは無理であるから、ほどよく胚の出たときに、スポイトで胚を吸上げてスライドグラス上に移し、細い筆を使って卵黄を除去する。スポイトの使い方、筆の用い方にも要領がある。
胚の発音階程の鑑別だけならば、慣れれば、卵黄をきれいに取去る必要はない。
卵黄を除去した胚は、そのまま、またはメチレソブリュー(0.1%ぐらいの水溶液)を滴下して適当に染色し、余分の色素液を濾紙で吸取って鏡検する。
水中に長くおくと胚が膨潤するから、スライドグラスに移してからは、30%アルコールを加えて、水と置き換える方がょい。メチレソブリューを30%アルコールに溶解しておき、染色と置換とを同時に行なうのも一つの方法である。しかし、アルコールは乾き易いから、長くおきたい場合にはグリセリンを1滴加えて、カバーグラスをかけておくと、透明になり、環節もよくみえ、数日間は保存できる。しかし、蚕種保護のために、発育段階を毎日調べる作業としては、簡単な解剖で即座に鑑別のできるように熟練することが必要である。このための顕微鏡は50倍以下の倍率がよく、慣れれば20-30倍ぐらいの低倍率の方が能率が上がる。
固定した卵を、直ちに解剖せず、後日の調査用に保存するためには、30%アルコール中に入れておく。
30%以上のアルコールでは、胚が硬化してよくない。固定の度が過ぎたり、アルコール中で硬化して胚の脆くなった卵は、塩酸アルコール(30%アルコール100ccに塩酸1ccを加える)に数分間浸してから解剖すると、胚が柔軟になり、解剖し易い。
発育段階を知るための胚の観察は抽出検査であるから、沢山の卵を乱暴に解剖して、その中から、きれいに取出せた少数のものだけを調べたのでは意味をなさない。材料にした卵は少数でも、30粒なら30粒全部の胚が検査できるように解剖しなければならない。卵の解剖は胚の発育段階によっても難易があるから、壊れないで残ったものだけによって判定すると、誤りを犯すことがある。
卵殻を除去する方法としては、針や解剖刀で一々切除せず、次亜塩素酸ナトリウムや(1166,1167)、苛性カリ(253,910)などを用いる方法も行なわれている。これらの方法は卵殻を薬品によって溶かすもので、多数の卵を短時間に処理できるから、能率がよいように思われるが、胚の破損も多く、注意しないと抽出検査の主旨にそわないことがある。
簡易解剖法、取出した胚を永久標本にする方法、卵全体を透明にして胚をみる方法などについては多くの文献がある(25,106,230,231,288,292,293,457,521,525,580,644,667,668,723,725,726,728,794,811,910,935,1054,1183)。
b 生体解剖法
生体解剖法は普通には用いられないが、事故のあった卵の生死の鑑定や、死因の追究などには役立つ方法である(925,932)。また、卵黄細胞の遊離性や卵黄細胞内における脂肪性顆粒の集合、分散Ⅵ2Ba,b)なども生体解剖によらなければ観察しにくい。慣れると生体解剖によって胚の発育段階の識別もできる。
生体のまま胚を取出すには、だるま形期(Stage5)以前のように胚が卵表の広い部分を覆っている時期には、卵の背側にそって、その中心線を切り開くのがよいが、それ以後の時期の卵においては、卵の中央、水引きの部分を通して長軸にそい、漿膜に達する深さで卵殻を切り破り、切りロの片側(楕円の半分)の卵殻を除去した後、生理食塩水(0.8%NaCl)中に沈め、解剖針の先きで漿膜に包まれた卵の内容をあやつるようにして卵殻から浮き出させる。次ぎに、浮き出た卵内容の漿膜の切り口に2本の針を入れて左右に開き、卵黄と胚とを漿膜から離す。体晩節においては、漿膜を開いても、その内面に卵黄に埋まった胚が付着していて、容易に離れないが、前休眠期および臨界期の卵においては、漿膜を切っただけで、こぼれるように卵黄細胞が出てきて、卵黄と胚とも離れ易い。卵黄細胞の遊離性と云うのはこれである。
最初、卵殻を切り破るときに、同時に漿膜を破っておかないと、あとで漿膜を開く操作がむずかしくなり、胚を損傷することが多い。固定した卵と違い、生体では、胚に弾力がある上に、卵内で移動し得るため、針で胚を突いても案外損傷が少なく、だるま形期以後には思ったよりも容易に完全な形で胚が取出せる。
卵黄核や分裂核を観察するには、このようにして取出した卵黄のかたまりをスポイトでスライドグラスの上に移し、0.05%ぐらいのメチレンブリュー(0.8%NaClに溶解)1滴を加え、カバーグラスをかけて観察する。卵黄細胞はカバーグラスの上から押さえると潰れるから、液を十分に加え、そっとかぶせただけで観察する。卵黄核をみる場合には卵黄細胞を潰した方がよい。
生体による観察は、生理食塩水中での胚や卵黄細胞の変形、核内および核中での微小顆粒の動き、凝集、分散、染色性の変化などを通じて卵の生理状態を考えようとするのであるから、細かい注意と熟練とが必要である。染色には、必要に応じて、メチレンブリューの外、各種の生体染色用の色素を使用する。
毛細管を用いて卵内から胚を吸出す方法もある(1161)。
B 孵化調査の注意
孵化調査と云えば、卵を催青し、孵化が始まれば毎日、機械的にその数をかぞえ、後で型通りの計算を行なえばすむように思われがちであるが、実際には、試験の目的に応じて考えなければならない多くの問題がある。
まず日常作業についての問題点から考える。試験用の催青は掃立て用の大量催青とは違い、狭い恒温室または恒温器の中で行なわれることが多い。温湿度の調節は狭い場所においても支障なく行なうことができるが、問題は照明である。狭い処に点燈すると器内の温度が上り過ぎる。この点を往々工夫しても、蚕種の量が多いと、台紙を1枚並べにすることができず、光のよく当ったものと蔭になったものとでは、孵化に数時間の遅速を生ずることがある。このような卵を取出して何気なく孵化卵をかぞえていると、最初に数えた区とあとでかぞえた区との孵化成績が非常に違う。また、調査中にも続々発蟻するものがあるが、これを羽箒で掃くために、孵化し損うものもできる。孵化歩合を集計してみて、最多2日孵化歩合の山のはっきりしていない場合の中には、活性化状態のほかに、このような不注意が原因になっていることもあるが、不慣れな臨時の補助者などにまかせ切りにしていると、毎日みていながら気付かないことが多い。孵化調査を始めるときには、その日に発蟻する分は全部出尽しているように条件を整える必要がある。
大量の孵化調査に追われているような場合には、切角温湿度調節のしてある催青室から蚕種を持出して、半日近くも実験室に放置していることもない訳ではない。これを毎日繰返えしていると孵化に影響がある。点催青卵は漿膜がなくなっているので乾き易く、湿度が低いと孵化かおくれたり(Ⅵ3Db)、催青死卵がふえたりするが、このような被害は、一斉に孵化する場合よりも孵化の不斉な場合に、早く孵化するものよりもおくれて孵化するものに多いから、孵化を益々悪くすることになる。更に問題なのは死卵と不受精卵との区別である。これは色が着いていないものは不受精卵、色の着いているものは死卵と云うように簡単に割切れるものではなく(ⅩⅠ1
)、熟練者がみても完全には識別できないものである。
このようにして得られた数字だけから、小数点下までの孵化歩合を計算し、僅かに1、2%の差で孵化の優劣をきめたり、統計的な有意性を論じたりすることは十分に注意しなければならない。
また、学校や試験場の試験においては、例えば、一方の区の孵化歩合は70%であり、他方は80%で、統計的にも有意な差であったから後者がよい、と云うような結論をしているのを見かけることがあるが、これにも問題がある。学問としてはそれでよいのであるが、蚕種保護の実用の面からみれば、孵化歩合80%では物の役に立たないから、どちらの区も悪いと云わなけれはならない。対照区と比較して良し悪しを論ずる90%以上の場合の差でなければ、その結果に興味を待ち得ないのである。
同じことは産卵数についても云える。産卵数増加のための飼育あるいは育成の結果が、基準以下の卵数の範囲で論じられても役に立たない。
孵化の斉否は最多2日孵化歩合(実用孵化歩合)によって示されるのが普通であるが、これにも問題がある。中間手入れによって胚の発育程度の揃えてある蚕種においては、この最多2日の時期は孵化が始まってから2、3日のうちであることが予測できるが、活性化の初期から始めて、種々な時期の蚕種の日別孵化歩合を調べ、これを曲線に描くと、活性化の進み具合によって、逆L字型、S字型、J字型などの曲線が得られると云う市川・剣持(272)の調査からもわかるように、自然温度て保護した蚕種においては、ただ最多2日孵化歩合を示されただけでは、必らずしも最多2日孵化の時期を知ることができない。これは越年種に限らず、冷蔵浸酸種などについても同様で、孵化の不斉になりがちな蚕種については、最多2日が催青何日目であるかを明示することが、2夜包み、3夜包みの問題と関連して、実用上必要なことである。
これらの点をどうするかは広く検討されなければならない問題であるが、実用に焦点を絞った孵化調査法として著者の試みているのが催青2週間孵化歩合、初発2日孵化歩合などである。
完全に活性化している越年種ならは、甲胚子の時期から始めても催青2週間で孵化するものと考え、催青を始めてから2週間目の孵化歩合を調べて活性化状態を知ろうとするのが前者である。2週間目の調査によって活性化の不十分な場合には、催青3週間または4週間の調査を行なう。これは催青15日、20日、30日などとしても差支えない。
初発2日孵化歩合は主として即時浸酸の場合に、最多2日孵化歩合の代りに用いる。即浸種は初発日とも2日間で大部分発蟻しなければ実用にならないと云う考えから、これによって浸酸処理の適否をみようとするものてある。催青を始めてから毎日蚕種の状態に注意し、初発蟻の出た日を確認して、その翌日に孵化調査をする。初発2日で孵化しなかった卵が死卵であるか否かをみるためには初発3日または5日の調査も行なうが、その必要のない場合には初発2日で火にあぶって未孵化卵を殺しておけば、その日の中に卵をかぞえなくてもよい。
初発3日孵化歩合は冷蔵浸酸種および越年種の孵化調査に用いる。しかし、初発3日では、催青の面からみると3夜包みの蟻蚕が入ることになる。3夜包みの蟻蚕は実用上好ましくないので(Ⅸ3C)、これらの蚕種についても、基準を初発2日に高めて、保護、取扱い法の改善に努める必要がある。初発4日以後の発蟻は実用上は無効発蟻と考えるべきものである。
これらの方法では毎日の孵化調査に追われることがないので、余裕をもって催青条件や卵の状態に注意することができ、孵化してしまえばわからなくなるような点催青の僅かな遅速を記録したり、初め催青2週間と予定していた調査日を繰上げ、または繰下げるなどの臨機の処置を行なって、試験区間の違いを明確につかむこともできる。
孵化卵数と廃蟻数とをかぞえ、その差を掃立て頭数とし、これを基準にしてその後の減蚕歩合を求めることは普通に行なわれるが、4令起蚕の頭数がこの掃立て回数より多くなっており、掃殼を再調査すると、最初の孵化卵数が間違いであったことの判明するような場合も絶無ではない。4令頭数が増加していた場合には再調査を行なうが、減少している場合には余程のことがなければ再調査は行なわないから、間違った掃立て蚕数がそのまま用いられていることがあるものと考えられる。これは、孵化卵にインクで印を付けてかぞえていた従来の方法の代わりに、蚕卵計数器(255)やこれを電気的な方法に改良した装置を使用すれば防止できるが、何時、何処ででも簡単に行なうことのできる従来の方法は今後も残るものと思われるから、卵数調査には注意が必要である。
実用上の孵化歩合の基準を95%以上、または90%以上にとり、これに満たないものは失格とするような場合には、催青何日または初発何日と云う調査法を採用すれば、目的によっては、目測で孵化調査を行なうこともでき、本多(214)の死卵調査器のようなものも使える。
台紙に産ませてある卵を試験区数に切り分けて用いる場合に、卵を傷つけると、付近の卵にその卵汁が付着して孵化を悪くするから注意を要する(65,834)。 従って、台紙は鋏で切るよりも引き裂く方がよいが、卵を強く摘まむと、潰れなくても孵化の悪いことがある。
バラ種の孵化調査は、卵を紙に貼り付けて行なうのが普通であるが、糊が多過ぎると孵化に影響がある。最も簡単なのはセロハンテープに卵を貼る方法である(879)。バラ種をセロハンテープの巾よりやや狭い長方形に拡げ(長さは適宜)、セロハンテープの粘着面を下に向けて卵面にかぶせ、上からなぜると卵はテープに貼り付く。テープの両端に余裕を残しておき、この部分を反対に折り曲げ、卵を貼った面を上にして厚紙に貼り付け、粘着面にはタンポンか綿塊でタルクを打ち付けておく。タルクを忘れると、取扱いに不便なばかりでなく、発蟻の途中で蟻がテープに粘着して孵化し得ない。テープは巾の広いものが便利である。
各蛾から切取った1片ずつを集めて1試験区とした場合に、その孵化歩合を1試験区全体の総卵数で出してあるのをみることがあるが、各片別に孵化歩合を求めて、統計的に処理するのがよい。なお、前記初発何日かの孵化歩合を求める場合には、各蛾別ではなく、その区のどの蛾区かに初発があれば、その日を初発日とする。
2連制の成績は平均ばかりでなく、別々に比較してみることも大切である。例えば、一方が95%、他方が65%で平均80%の場合と、両方とも80%で平均が80%の場合とは、その意味が同じとは云われない。このようなことからも重要な問題に気付くことがある。
Ⅷ 人工孵化
人工孵化法には種々あるが、実用になっているのは浸酸法だけであると云ってよいから、主としてこれについて述べる。その他にどのような方法が試験されたかについては、石森(258a)、三浦(547)、田中(1036)などを参照されたい。
日本においては、現在、春蚕よりも夏秋蚕の比重が大きいが、この夏秋蚕を支えているのは人工孵化法-浸酸法であるから、その方法が蚕糸業の中で果している役割は非常に大きい。しかし、これだけ普及した浸酸法がとのような働きで越年性卵を不越年化させるのかは殆どわかっていない。
三浦(553)は、塩酸のCl-イオンが卵に吸収されて効果を発揮するものと考えているが、吸収されたCl-イオンの働く機構については言及していない。三浦の考えは、人工孵化可能な時期の蚕卵をガラス管に入れ、これに一方の電極を挿入し、他方の電極は裸のままとして、共に塩酸の中に浸すと、卵を入れた方が負になって電流が流れると云う実験結果に基ずいているのであるが、四方(835)の追試によれば、この実験結果には問題がある。上原(1092)は、塩酸中に両極を入れ、その各々の付近で浸酸を行ない、また食塩水中においても同様な試験を行なったが、Clイオンの働きと浸酸の効果とを直接に結び付けるような結果は得られなかった。
1 浸酸成績に影響する一般的な条件
A 蚕卵の条件
上蔟後の保護温度によって浸酸有効浸漬時間の巾が相違するが(Ⅲ3Bdii)、産卵後の保護温度は浸酸適期の巾に影響する(第122表)。
| 産卵後の日数 | 保護温度 | ||
| 25℃ | 20℃ | 15℃ | |
| 日 時間 10 18 1・ 0 1・ 8 1・16 2・ 0 2・ 8 2・16 3・ 0 3・ 8 3・16 4・ 0 5・ 0 6・ 0 7・ 0 8・ 0 9・ 0 10・ 0 11・ 0 |
% 37 96 97 97 93 95 81 63 5 9 2 3 2 0 0 0 0 0 0 |
% 31 93 96 97 94 97 94 97 91 82 38 10 19 0 0 0 0 0 0 |
% 20 45 6 89 91 95 94 97 96 96 94 92 95 95 55 39 17 3 1 |
渡辺(1149)は第122表の例をも含めて3回の試験を行ない、その結果から、即時浸酸の適期の中心は、産卵後の保護温度が25℃の場合には、大約1日目、20℃の場合には2日目、15℃においては4日目であり、90%以上の孵化歩合の得られる期間の巾は温度が低いほど広いと結論した。勿論これは品種によっても異なるので、この数字がそのまま他の場合にも当てはまる訳ではないが、一般的な傾向としてはこの通りである。
冷蔵浸酸の場合の、産卵後冷蔵までの保護温度と冷蔵時期との間にも同様な関係があって、保護温度の低い場合に冷蔵適期の巾が広い(第123表)。
| 冷蔵までの日数 | 保護温度 | ||
| 25℃ | 20℃ | 15℃ | |
| 日 時間 1・ 0 1・12 2・ 0 2・12 3・ 0 3・12 4・ 0 4・12 5・ 0 6・ 0 7・ 0 10・ 0 15・ 0 20・ 0 |
% 72 95 96 97 98 95 97 96 79 70 44 45 71 53 |
% 20 89 94 98 96 96 97 98 96 97 90 90 94 79 |
% 0 57 75 95 98 97 98 98 98 98 98 98 97 97 |
産卵後の保護温度によって浸酸および冷蔵の適期の巾にこのような違いのあるのは、胚の発育および越年化の進行が温度によって相違するためである。第122表および第123表において、早い時期の浸酸および冷蔵によって孵化の悪いのは、時期が早いと越年性卵が不越年化しにくいのではなく、発育がある程度進まなければ卵が浸酸に耐え得ずに死卵になるためであって、卵をいためることの少ない常温浸酸によれば、産卵後10時間以内の卵でも不越年化する(Ⅷ2B)。温度が高いと胚の発育が速く、浸酸に耐え得る時期に達するのは早いが、同時に越年化の進行も速いために適期の巾が狭いのである。
産卵後の時間が浸酸成績に影響することも、浸酸成績の向上には短時間内にまとめて産卵させ、卵令を揃える必要のあることも、これによって理解される。
小形な卵は大形な卵に比べて浸酸刺戟に対する抵抗力が弱い。
大村(753)によれば、4令2日目に片側の卵巣を除去すると、残りの卵巣の卵が大きくなり(Ⅰ2Abi)、また上蔟のときに尾角を切って出血させた蚕の卵は小さいが、産卵後20時間目に比重1.075、46℃の塩酸で4分および7分の浸酸を行なって比較すると(日112号×支110号)、4分間浸漬においては卵の大小と孵化歩合の順位との関係は顕著でなかったが、7分間浸漬においては卵の大小(片側卵巣の卵>無処埋蚕の卵>出血蚕の卵)と孵化歩合とが明かに比例し、また、小卵ほど最多2日孵化日がおくれた。蛹が大きいと小さいものに比べて一般に卵が大きく、その卵の大小が浸酸成績に影響する(572)。
橋爪(174)によると、全令を桑で飼育した蚕と柘で飼育した蚕との卵は大きさが異なり、支11号についての実験によれば、100粒平均の長径×短径が前者1.34mm2、後者1.19mm2 、1g卵数は前者1,821粒、後者2,139粒であったが、この卵を浸酸した成績を第124表に示す。
| 浸漬時間 (分) |
区別 | 孵化歩合 (%) |
日別孵化歩合(%) | 催青死卵 (%) |
不受精卵 (%) |
|||
| 第1日 | 第2日 | 第3日 | 第4-7日 | |||||
| 3 | 桑飼育(大卵) 柘飼育(小卵) |
82 93 |
56 57 |
17 36 |
9 4 |
17 1 |
17 6 |
3 6 |
| 5 | 桑飼育(大卵) 柘飼育(小卵) |
93 92 |
79 49 |
16 42 |
3 6 |
1 1 |
6 7 |
3 6 |
この成績において、3分間浸漬の場合に桑飼育の孵化歩合が劣り、孵化も不斉なのは浸酸刺戟が不足なためであり、5分間浸酸で柘飼育の孵化歩合が劣り、孵化不斉なのは浸酸刺戟の強過ぎたためであって、柘飼育の蚕の卵が桑飼育のものに比べて浸酸刺戟に敏感なことを示している。これは22時までの産卵についての成績であるが、22時以後の産卵(卵令が若く卵も小さい筈である)を上と同時に浸酸した結果では、浸酸時間4分、7分ともに桑飼育の成績が勝っていた。
橋爪はまた、片側卵巣摘出および幼虫の出血によって作った大卵および小卵についての浸酸成績も比較しているが(明記していないが桑飼育と思われる)、14-22時の産卵を翌日16時に比重1.075、液温46℃の塩酸で浸酸した結果では、大卵区は浸酸2分間では孵化歩合が低く、孵化不斉、3分間でもなお刺戟不十分であった。対照蚕の卵は2分間でも可なりの成績を示し、3分間では更によかったが、幾分刺戟不足の気味があった。これに対して、小卵区の卵は浸酸2分間でほぼ十分であり、3分間ては強過ぎるぐらいであった。
卵の大小によって浸酸刺戟に対する感受性の異なる理由としては、小卵は体積の割合に表面積の大きいこと、卵殻が薄く、塩酸が通り易いのであろうと云うことなとが考えられるが、その証明はない。
塩酸に対する卵殻の透過性は蚕品種によって相違がある(916,937)。
浸酸刺戟に対する蚕卵の感受性は蚕品種によって著しく相違するが、福田・河野(127)は淘汰によって、即時浸酸で孵化し易い系統と孵化しにくい系統とを分離し、冷蔵浸酸についても、冷蔵期間15日で容易に孵化する系統と孵化不良の系統とを分離した。
B 塩酸の条件
a 夾雑物
現在、日本で作られている塩酸は大部分が合成法によるものであるから、夾雑物について特に心配することはないが、夾雑物の種類および含有量によっては蚕卵に害がある。
夾雑物としては硫酸、硝酸、亜硝酸、遊離塩素、砒素なとが挙けられているが、塩酸の製造法によって異なるので一概には云えない。新しく購入した塩酸は予備浸酸をして、蚕卵に害のないことを確めてから使うのが安全である。
夾雑物についての大櫛・地引(730)の試験結果は次ぎの通りである。
硫酸 普通の工業用塩酸に含まれる程度の濃度では蚕卵に害はない。比重を調整した後の浸酸用塩酸液に硫酸2.5%以上を加えて浸酸を行なうと孵化が悪かったが、これは硫酸を加えた分だけ成が濃くなっているためで、硫酸を加えた後に比重を調整した場合には被害がなかった。
亜硫酸 比重を調整した塩酸液に0.05%以上の亜硫酸を加えて浸酸を行なうと、卵は殆と全部死卵になる。しかし、亜硫酸は温度を上げると揮発するから、開放状態で加熱、撹拌しながら行なう普通の浸酸においては、被害は殆どないものと考えられる。上記の実験は密閉状態で行なったものである。新らしい塩酸はよく加熱、撹拌した後に使うのがよい。
硝酸 ホルマリンを加えない場合には、比重を調整した塩酸液に2.5%の硝酸を加えても蚕卵に被害がなかった。 5%加えると著しい被害があった。普通の工業用塩酸に夾雑物として含まれる硝酸の量は、硫酸よりも少ないと考えられるから実際上の心配はない。
ただ、塩酸にホルマリンを混ぜると、これが硝酸に作用して蟻酸と亜硝酸とを生じ、亜硝酸は無水亜硝酸に変り、更に酸化窒素と二酸化窒素とになる。この反応は低温でも起こるが、温度が高いと益々促進される。反応中は泡が発生し、熱を発し、液が褐色を呈するが、反応が終了すると無色になる。
比重を調整した、ホルマリンを含む(100cc中にホルムアルデヒド0.5g)塩酸中に硝酸を加え、上記の反応の進行中に蚕種を投入して浸酸を行なうと、硝酸が0.01%の場合には被害がなかったが、0.1%のときは全部死卵になった。しかし、反応終了後に浸酸すると、硝酸を1%添加した区においても被害はなかった。
このように、ホルムアルデヒドは硝酸と反応して亜硝酸を生ずるが、この反応は、硝酸1gに対してホルムアルデヒド約0.5gの割合で行なわれるから、蚕卵の脱落防止のために塩酸液にホルマリンを加える場合には、まず夾雑物として存在する硝酸の約半量に相当するホルムアルデヒドを加え、加熱して反応を行なわせ、反応終了後に、脱卵防止に必要な量のホルムアルデヒドを加えればよい。蚕卵にホルマリンを塗布し、新らしい塩酸を用いて浸酸を行なう場合には、塗布したホルマリンを乾かしてから浸漬するのが安全であり、また脱卵防止の効果も確実になる。
亜硝酸 0.01%までは塩酸液に加えても害がなかったが、0.025%加えると孵化歩合が低下し、0.05%では殆ど全部死卵になった。しかし、亜硝酸は熱に対して不安定であるから、この害のあった塩酸を用いて、1週間後に行なった浸酸においては全然被害がなかった。
即ち、普通の浸酸を行なうのと同じ温度に塩酸液を加熱して、数分間撹拌すれば、亜硝酸含量0.15%以下の場合には無害になるものと考えられる(0.15%以上の場合は実験していない)。但し、亜硝酸は不安定で、前記のように無水亜硝酸、酸化窒素、二酸化窒素を生ずるから、直接に有害なのがどれであるかは不明である、兵頭(232)は過酸化窒素(二酸化窒素)が有害であるとしている。
砒素 0.01%までは害がなかったが、0.025%では孵化歩合が可なり低下した。
塩化鉄 塩化第一鉄(FeCl2)を塩酸液に5%添加しても無害であった。
土釜を用いて浸酸液を加熱すると、土釜に含まれている塩化鉄が塩酸内に溶出して、卵殻を着色させることがある。清水ら(854-856)は土釜から溶出した塩化第二鉄(FeCl3)によって塩酸液が橙青色を呈し、これで浸酸した卵の卵殻が暗緑色に着色した例を報告しているが、孵化歩合には影響がなかった。塩酸液に塩化第二鉄、塩化コバルト、塩化第一銅、塩化マンガン、塩化亜鉛などを1%添加して浸酸しても、孵化および蟻蚕の絶食生命時数に影響がなく、飼育試験の結果からみても無害であった。これらの中で卵殻を着色させたのは、塩化第二鉄のほかは塩化第一銅(CuCl)のみであった(黄色)。塩化第二鉄および塩化第一銅による卵殻の着色性は蚕の系統および品種によって相違があると云う。
塩化バリウム 1%以下の添加では無害であった。
以上の結果からみると、普通の夾雑物のうち、硫酸、亜硫酸、硝酸なでにはそれぞれ対策があり、塩化鉄、塩化バリウムは無害で、何れも心配することはない。注意しなければならないのは0.01%以上の砒素だけである。
遊離塩素 は極めて有害であるが、非常に揮発し易く、実害は殆どないと思われる。なお、普通の意味の夾雑物ではないが、浸酸中に寒暖計がこわれて、塩酸液に水銀が混ざるようなことがあっても、蚕卵には被害はない(226a,383a)。
b 濃度の調整
純粋な塩酸であれば、その比重によって塩化水素(HCl)の濃度を知ることができる。第125表は15℃、真空空中で測った不純物を含まない塩酸の比重と塩化水素の濃度との関係を示すものであるが、実用上は、真空中と云う条件をはなれ、普通の純度の塩酸に対してこの表を適用する。
| d415 (真空中) |
HCl (%) |
d415 (真空中) |
HCl (%) |
| 1.000 1.005(16) 1.010 1.015 1.020(21) 1.025 1.030(24) 1.035 1.040(27) 1.045 1.050(30) 1.055 1.060(33) 1.065 1.070(36) 1.075 1.080(39) 1.085 1.090(42) 1.095 1.100(45) 1.105 1.110(48) |
0.16 1.15 2.14 3.12 4.13 5.15 6.15 7.15 8.16 9.16 10.17 11.18 12.19 13.19 14.17 15.16 16.15 17.13 18.11 19.06 21.01 20.97 21.92 |
1.115 1.120(51) 1.125 1.130(54) 1.135 1.140 1.1425(57) 1.145 1.150 1.152(66) 1.155 1.160 1.163 1.165 1.170 1.171 1.175 1.180 1.185 1.190 1.195 1.200 |
22.86 23.82 24.78 25.75 26.70 27.66 28.14 28.61 29.57 29.95 30.55 31.52 32.10 32.49 33.46 33.65 34.42 35.39 36.31 37.23 38.16 39.11 |
比重と塩酸濃度との関係は温度によって相違するから、15℃以外の温度において測定した比重は補正しなければならない。このためには、温度1℃の上下によって比重がどれだけ変化するかと云う補正値を求めておき、これを加除する方法(233,389,580)や、塩酸の膨張係数を用いて計算、作成した表を用いる方法(603,622)などがある。
第125表の括弧内に示した数字は温度1℃の上下による比重の補正値(×10-5)を示したもので、例えば、15℃における比重1.070の塩酸は温度が1℃上れば36/100,000=0.00036減少して1.070-0.00036=1.06964になり、1℃下れば1.070+0.00036=1.07036になることを示す。15℃比重1.075の場合の補正値は1.070の場合の36と1.080の場合の39との平均によって37.5×10-5が得られる。表の脚註に、補正値の適用範囲は10℃- 20℃であると注意されてはいるが、この補正値をそのまま31℃の差のある46℃に適用して計算すると、46℃での比重は
1.075-37.5×10-5×(46-15)=1.063375
になる。
小針(389)、水野(580)の補正値は1℃の上下につき
13-19%の塩酸については 約0.0003
20-27% 〃 約0.0004
28-35% 〃 約0.0005
となっているから、それによって計算すると、15℃比重1.075の塩酸は46℃においては比重1.0657になる。
室賀(622)の表によれば同じ比重が1.0642である。
このように、それぞれの値が僅かに相違しているが、普通の比重計では読みとり難い違いであって、何れの方法によって比重を調整しても人工孵化施行の実際問題としては差支えない。
15℃比重の1.10および1.110は48℃において、第125表の計算によれば1.08515および1.0932、小針らの補正値によれば1.0868および1.0968、室賀の表によれば、1.0853および1.0943である。
塩酸濃度について重要なのは、これよりも比重について違った解釈があり、“塩酸の液温46℃、比重1.075”と記されている場合に、これを“15℃において比重1.075の塩酸を46℃に温める”と解釈する人々と、“46℃に温めたときの比重1.075”であるとする人々とがあって、誤解の基になっていることである。
古くは、15℃比重て塩酸濃度を調整し、これを目的温度に温めて使用するのが普通てあったが(58,389,411,580)、その後、塩酸液を46℃に温めてから比重計を浮べて1.075に調整し、使用することが行なわれるようになり、現在、蚕種業者の約半数はこの方法によって浸酸を行なっているものと思われる。農林省蚕糸試験場の内部においてさえ、この二つの慣行があって、共通試験の規定をそれぞれに解釈していることがある。
同じく比重1.075とは云っても、46℃での1.075の塩酸は15℃比重のものに比べて、塩化水素の濃度としては約2%高い。どちらの比重がよいかは別問題にしても、もともと15℃比重による試験の結果採用された1.075を温度の異なる場合にそのまま適用すると結果の相違するのは当然である。
自分の処だけで一つの目安として使っている限りでは、どちらを用いても差支えないが、日本中、あるいは世界を通じて話し合う場合や、他の試験成績を参考にする場合などには、誤解や事故のもとになるおそれが多い。
15℃比重1.10の塩酸を用いる限り、冷蔵30日間の冷蔵浸酸において卵の潰れることは殆どないが、目的温度47.8℃での比重1.10の塩酸で浸酸すると卵の潰れる危険がある。従って、発表した試験成績や講習会で話したことが思わぬ事故を招く心配がある。
このような混乱の起こったのは、渡辺の報告(1140,1149)が、総べて、目的温度での比重によって塩酸濃度を示していることからわかるように、浸酸液調製上の便宜から、当時の蚕業試験場においてはこのような指導が行なわれていたためであろうと思われるが、渡辺自身(1149)は、第127表に示す成績その他から、“即時塩酸孵化法においては、塩酸液の濃度は液温46℃にて比重1.06又は1.065となすを有利とすべく、此濃度は略春蚕採種期の室温にて比重1.075となし後之を46℃に昇温せしめたる場合に相当する”と述べ、目的温度での比重1.075は濃度が高過ぎると注意している。中井(649)も、塩酸比重は15℃できめるのが学術的であるが、実際には夏季の天然温度できめて差支えない。しかし、このようにして調製した塩酸液を115゚F(46.1℃)に温めたときにもなお1.075を維持しなければならないと考えて塩酸を補充すると死卵を出す原因になる、と云う意味のことを記述している。これらの真意の理解されなかったことが混乱の原因になったものと考えられる。
このような現状においては、比重には必らず調整したときの温度を付記する必要がある。本書に引用した多くの浸酸成績の中にもこの点の不明なものが少なくなかったが、わかる限りは15℃比重、46℃比重と云うように明記した。
15℃比重を用いると次ぎのような利点がある。
1)購入した塩酸に表記されている比重は15℃比重であるから、第125表によって濃度がわかるが、表を用いないでも、15℃比重の値から1を引き、小数点下だけに200を掛ければ概略の濃度(%)が求められる。即ち
(15℃比重-1)×200=塩酸の概略濃度(%)
2)このようにして原液濃度(%)がわかれば、次ぎの式を用いて、原液を目的濃度(%)に稀釈することができる。目的濃度(%)に稀釈するために原液1㍑に混合すべき水量(x)は
{原液濃度(%)-目的濃度(%)}/目的濃度(%)=x(㍑)
3)比重によって原液を稀釈する場合に、目的比重の塩酸100㍑を作るのに要する原液の量(x)は
(目的比重-1)/(原液比重-1)×100=x(㍑)
このような計算によって概略の稀釈をした後、比重計を浮べて細かい調整を行なうのである。
c 浸酸による塩酸濃度の変化
何回も浸酸に使用した塩酸でも、比重さえ調整すれば、新らしい塩酸と同様に使用できる場合もあるが(529,531)、比重の調整だけでは不十分な場合も少なくない。水が加わって、濃度と共に比重の下がったような場合には、濃度の低下が比重の減少として現われるから、塩酸を加えて比重を調整すればよいのであるが、卵の膠着物質やバラ種台紙の糊などが多量に溶解すると、塩酸濃度は下がっていても、比重はその割に減少しないことがある(944)。
新らしい塩酸は蚕卵に害があるとして、前年または前蚕期に使用した古い塩酸を、新しい塩酸に半分以上も混合して浸酸用塩酸液を調製する習慣が広く行なわれているが、このような習慣のあるのは、上に述べたような目的温度における比重調整を行なっている所が大部分である。従って、古い塩酸に特別の効果があると云うよりも(極く一部には、先きに述べた有害夾雑物の稀釈に役立っている場合があるかも知れないが)、混入物のために比重の割には濃度の低くなった塩酸を混合して、塩酸濃度の高過ぎるための被害を意識せずに軽減しているもののように考えられる。これは、これらの場所で使用している浸酸用塩酸液を調べてみると、目的温度で比重を調整しているにも拘らず、塩酸濃度(%)としては、目的温度での比重に相当する濃度と15℃比重に相当する濃度との中間ぐらいに下がっている場合の多いことが示している。
15℃比重による浸酸の共通試験を行なった処、ある特定の場所での孵化が毎回悪く、塩酸濃度の低いためのように考えられる場合があった。原因を調査した結果、ここでは、従来、古い塩酸を混合して、目的温度で比重を調整する習慣があったので、15℃比重の場合にも古い塩酸を混ぜたため、比重は同じでも所要の塩酸濃度(%)の得られなかったことがわかった。即ち、古い塩酸を混合することは濃度の高過ぎる場合には効果があるが、もともと濃度の低い場合には効果がなく、却って浸酸効果を低下させることさえある。技術はこのように関連し合った一連の条件の上で成立するもので、その一部分だけを、前後に関係なく取り換えても効果の挙がらないことに注意しなければならない。
古い塩酸の混合には、このように有効な面はあるが、古さの程度(比重と濃度との関係)が常に同じとは限らないため、同じ量の古塩酸を混合し、その比重を同じに調整しても、できた浸酸用塩酸の濃度(%)の一定しない点に問題がある。
濃度を下げて蚕種の被害を防止するためだけならば、このような不安定な方法よりも、新らしい塩酸を用いて初めから低い濃度で使用すれば十分足りる訳であるが、経済的な理由もあって古い塩酸を一部使用するのであれは、比重以外の方法によって塩酸濃度を確かめる必要がある。最初の濃度をいくら正確にしておいても、使用中に変動するのであるから同じであると云う考えもあるようであるが、浸酸の現場をみると、秒時計を使い、浸漬時間には非常に神経を使っている。秒をあらそうほど正確な浸漬を行なっても、液の濃度が最初から不定では効果の安定は望めない。
塩酸比重ぞ示す場合には、温度ばかりでなく、古い塩酸を混ぜたか否かも記す必要がある。
比重には出ない浸酸用塩酸液の濃度変化を、現場で簡易に検定する方法の一つの試みとしては次ぎのものがある(944)。
浸酸用塩酸液濃度の変動許容範囲を15℃比重±0.002 (以下、上限を+0.002下限を-0.002と記す)に対応する濃度(%)とし、この上限および下限濃度の塩酸液1cc によってそれぞれ中和する炭酸水素カリ(KHCO3)液を作りこれにメチルオレンジ液1滴を加えたものを検定液とする。検定液はポリエチレンの袋またはアンプルに封入して準備しておく。
濃度を調べたい塩酸液1ccをピペットで正確にとり、試薬中に入れて浸盪すると、検定液が黄色のままで変色しない場合、赤変する場合および橙色を呈する場合がある。この色によって塩酸液の濃度が上記許容範囲内であるか否かが判定できる(第126表)。
| 類別 | 検定液の種類 | 呈色 | 結果の判定 |
| a | +0.002 -0.002 |
黄 黄 |
塩酸濃度が許容範囲より低い |
| b | +0.002 -0.002 |
赤 赤 |
〃 〃 高い |
| c | +0.002 -0.002 |
黄 赤 |
〃 〃 内である |
この場合、厳格に云えば、膨張係数の関係から、被検塩酸を15℃に冷やしてからその1ccをとらなければならない訳であるが、実際問題としては、浸酸に使用中の温度のままの塩酸を用いて差支えない。なお、蛋白その他の溶解している塩酸液は、塩酸濃度は等しくても、卵に対する浸酸効果が等しいかどうかの問題もあるが、比重だけに頼るよりは浸酸成績が向上する。
検定液調製の1例を挙げると、15℃比重1.075に対する上限(+0.002)および下限(-0.002)濃度の塩酸は規定度がそれぞれ4.596および4.346であるから、その1ccを中和する炭酸水素カリとして、上限用には46.0172g/㍑下限用には43.5101g/㍑の水溶液を作り、これらの10cc宛にメチルオレンジ液1滴を加えたものを検定液とする。
C 塩酸液の濃度と浸漬時間との関係
塩酸液の温度が等しい場合には、濃度の高いほど適当な浸漬時間の巾が狭い(第127表)。
| 浸漬時間 | 塩酸比重 | |||
| 1.060 | 1.065 | 1.070 | 1.075 | |
| 1分 2 3 5 7 10 13 15 20 25 30 |
94% 94 97 98 97 95 96 95 96 91 86 |
97% 97 94 97 97 90 96 95 94 86 81 |
96% 98 96 98 97 95 95 91 83 61 54 |
96% 97 97 98 94 94 95 92 84 54 4 |
水田・佐藤(586)によれば、90%以上の孵化歩合の得られる有効浸漬時間の巾は塩酸液の濃度の高いほと狭いが、その中心値の変化は15℃比重の1.040から1.120までの間は直線的でそれ以上濃度が高まると急に緩かになる(第128表)。
| 比重 | 90%以上の孵化歩合の得られた浸漬時間 | |
| 中心値 | 巾 | |
| 1.04 1.06 1.08 1.10 1.12 1.14 1.16 1.18 1.20 |
396分 132 42 12 3.75 1.67 0.92 0.50 0.42 |
120-720分 36-228 12-72 4-20 2.5-5 80-120秒 50-60 30 25 |
水田らは、この現象を蚕卵が塩酸に反応するまでにある一定の時間が必要なためではないかと考えている。
D 塩酸液の温度と浸漬時間との関係
塩酸液の濃度が一定の場合には、液温の高いほど適当な浸漬時間の巾が侠い(第129表)。
| 塩酸の温度 | 浸漬時間 | 孵化歩合(%) | 最多2日孵化歩合*(%) |
| 80゚F (26.7℃) |
1時間 2 4 6 8 10 |
33 98 99 98 97 96 |
55 59 59 80 73 85 |
| 90゚F (32.2℃) |
1 2 2.5 3 4 5 |
98 97 94 97 97 88 |
81 87 93 86 90 91 |
| 100゚F (37.8℃) |
30分 1時間 2 2.5 |
98 93 95 99 |
90 97 97 91 |
| 110゚F (43.3℃) |
4分 16 32 40 |
93 94 95 90 |
88 97 94 97 |
| 120゚F (48.9℃) |
1 2 4 5 6 8 10 |
88 92 93 71 62 23 1 |
91 83 94 83 90 79 100 |
水田・苫米地(590)は、産卵時間を3時間に限った(支115号×支124号)×(日122号×日124号)の卵を、その中心時間から18-23時間目に15℃比重1.100の塩酸液に浸漬し、液温を0℃-50℃の間で変化させて試験を行なった(第130表)。
これによると、0℃-45℃の間では、液温5℃の上昇に連れて有効浸漬時間の中心値がほほ半分になっているが、45℃-50℃の間では更に急激に変化している。
| 塩酸の温度 (℃) |
90%以上の孵化歩合の得られた浸漬時間 | |
| 中心値(分) | 巾(分) | |
| 0 5 10 15 20 25 30 35 40 45 47.5 50 |
3,240 1,680 780 390 218 115 52.5 24 12 5.75 2.5 1.00 |
2,160 - 4,320 720 - 2,640 240 - 1,320 120 - 660 75 - 360 40 - 260 20 - 100 8 - 40 4 - 20 2.0 - 9.5 1.0 - 4.0 0.25 - 1.5 |
水田・高橋(587)によれば、15℃比重1.100および1.130の塩酸液を用い、液温5℃と-5℃とで即時浸酸成績を比較すると、この濃度差は、浸酸効果において、5℃の場合には大約浸漬時間12時間、-5℃の場合には同じく65時間の違いに相当する。材料は(日122号×日124号)×(支115号×支124号)であった。また、90%以上の孵化歩合の得られる浸酸時間は、液温が-5℃から+5℃に上昇すると15℃比重1.100の場合には約1/5、1.130の場合には1/2-1/3になった。比重1.100の場合に、10℃の液温上昇によって有効浸漬時間が1/5になるものとすると、液温25℃においては36分、37℃においては約6分になるから、従来知られている常温浸酸法および比重1.100の加温浸酸法の浸漬時間にほぼ一致すると云う。
塩酸液の濃度が1゚F下がれば浸漬時間を標準の1.2倍にし、1゚F上がれば0.83倍にするのがよいと云われているのは、液温10゚Fの上下によって有効浸漬時間が約5分の1または5倍になると考えての計算によるものである。
10゚Fは5.56℃であるから、1℃の上下に換算するには、
x5,56=5
を解いて x=1.34を得るから、液温が1℃下がれば浸漬時間を約1.34倍、1℃高いときは約0.75倍に伸縮することになる。
2 即時浸酸
即時浸酸は、蚕種に越年性の特徴が明瞭になる以前、または特徴の現われ初めた頃に浸酸を施して、越年化を中止させる方法である。浸酸後の有効冷蔵期間を25日、催青期間を10日とすれば、大約、産卵後10日以後35日までの間の掃立用蚕種を供給する入工孵化法である。加温法と常温法とに大別できる。
A 加温即時浸酸
a 浸酸
加温即時浸酸は、産卵後25℃、20時間目頃に行なうのが普通である。この時期を胚の解剖によって決定することはむずかしいから、産卵後の保護温度を一定にして、その時間によって処理時期をきめるのがよい。胚をみるには、解剖よりも、3%内外のホルムアルデヒド液(ホルマリン10倍液)を80℃に温めた中に蚕種を入れて5分間固定した後、卵殻の着いたまま60%アルコールに移し、10倍ぐらいの拡大鏡で観察するのが簡単で(1182,1183)、卵殻を通し、第24図Stage4のような胚の輪廓を透視することができる。
浸酸時期についての一般的注意は既に述べたが(Ⅷ1A)、胚がダルマ形(Stage5)に近付くほど、強い塩酸刺戟が必要になる(571)。
高島ら(966)は、日124号、日126号、日502号、支124号、支125号および支126号につき、産卵後、25℃で17±1、20±1および24±1時間目に15℃比重1.075、46℃、3、5、7分間の浸漬によって孵化成績を比較した。品種によって適期の巾に広狭があり、またその中心もずれていたが、総べてに共通する、浸酸適期は産卵後20時間目であった。
浸酸と云えば、蚕種を塩酸液に浸漬する操作に異常な注意の払われるのが常であるが、これまでに述べてきたように、浸酸成績に影響する条件は非常に多いから、浸漬だけにどのように注意を払っても、それだけでは浸酸成績を向上させることはむずかしい。
総べての条件を均一にしたつもりでも、結果は必ずしも同じにはならない。例えば、蚕糸試験場の本場および3原蚕種製造所において行なった共通試験成績をみても(第131表)各場所それぞれに、同じ掃立口ではあるが、産卵日の異なる材料によって試験を繰返えすと、浸酸時間3、5、7分の間の成績の相違よりも、同じ浸漬時間で浸酸日の違う区間の成績の違いの方の大きいことが珍らしくない。同じ3分とは云っても、毎回の浸漬時間には誤差があって、それがこの原因の一部になっていないとは云われないが、浸漬時間だけでは成績のきまらないことも明かである。
| 蚕品種 | 浸漬 時間(分) |
新庄 | 本場 | 小淵沢 | 宮崎 | ||||||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅰ | Ⅱ | Ⅰ | Ⅱ | ||
| 日124号 | 3 5 7 |
95 96 97 |
91 92 93 |
98 97 95 |
94 94 95 |
95 95 95 |
95 97 95 |
|
|
|
|
|
|
| 日126号 | 3 5 7 |
86 86 87 |
91 93 87 |
93 90 92 |
91 93 93 |
95 95 95 |
94 96 95 |
96 94 96 |
|
93 94 94 |
93 96 95 |
95 95 95 |
94 94 90 |
| 日502号 | 3 5 7 |
|
|
|
|
98 98 97 |
99 98 97 |
95 97 97 |
|
88 98 98 |
96 96 96 |
97 97 97 |
97 96 97 |
| 支124号 | 3 5 7 |
97 95 97 |
94 94 95 |
98 98 98 |
95 97 95 |
99 99 99 |
99 99 99 |
93 96 98 |
|
|
|
|
|
| 支125号 | 3 5 7 |
96 96 95 |
94 96 94 |
96 93 93 |
94 94 93 |
97 97 98 |
96 96 96 |
98 96 96 |
93 93 88 |
92 95 94 |
92 96 96 |
98 98 98 |
97 97 97 |
| 支126号 | 3 5 7 |
96 96 96 |
97 97 97 |
97 97 97 |
97 97 98 |
98 98 98 |
84 87 87 |
91 89 91 |
|
94 96 97 |
95 96 97 |
98 98 98 |
99 98 98 |
浸漬時間の一般的な標準は
日本種または日本種に近いもの 5-6分
支那種 〃 支那種 〃 4-5分
欧州種 〃 欧州種 〃 6-7分
交雑種は母体に準ずる。但し、塩酸濃度15%(15℃比重1.075、46℃比重1.064)、液温46℃の場合で、一化性は、二化性よりもやや長目の浸漬をする。液温に0.5℃以上の上下のないように注意する。
洗落したバラ種は水気を含んでいるので、そのまま浸漬すると塩酸液が稀薄になるばかりでなく、液温も下るから、浸酸用の本液の外に、これと同じ濃度で液温40℃ぐらいの予備液を作っておき、これに数十秒間浸した後、本液に入れるのがよい。
浸酸刺戟に対して非常に弱い品種がある。日122号が出た当時にこれが問題になって、種々な試験が行なわれた。
二木(143,144)によれば、日122号は産卵が揃わず、若い卵が混じていて、これが浸酸によって潰れる傾向があり、特に温度が低いと産卵がおくれる。
20℃においては25℃におけるよりも産卵最盛期が5時間ぐらいおくれるから、産卵中の保護温度を25℃に保つこと、20℃保護の場合には浸酸時刻をおくらせることが必要であると云う。
菅生(880)は、日122号の浸漬時間は、産卵後20-22時間では2分30秒-3分間、24時間では4分30秒がよいとした(15℃比重1.075、46℃)。
田中・神保(1029)は、浸酸による日122号の潰れ卵は、割愛後0-8時間、8-17時間、および17-22時間の卵を区別して浸酸すると、この順序で産卵のおそいものほど多く、若上げ蚕の卵、小形の卵、小さい蛹の卵および浸酸前冷蔵(5℃)を行なった卵にも多かった。蛹の出血および複眼着色期から触角着色期までの変温保護(午前9時-午後9時を30℃、午後9時-午前9時を18℃)も同じく日122号の浸酸による潰れ卵を多くした。
日122号については、また産卵後25℃に保護したものは20-25時間目に比重1.069、115゚F(46.1℃)で5-7分間浸漬するのがよいと云う報告(1126)、46℃比重1.075の塩酸液での浸酸は15℃比重のものに劣り、特に若上げ蚕の卵や卵令の若い卵にその影響の大きいこと(1071)、46℃比重と15℃比重との浸酸は胚の初期発育にも差を生ずること(934)、および卵殻の塩酸に対する透過性が大きい(916,937)、などの報告もある。
早春採りの蚕種は浸酸のむずかしい場合があるが、日122号においてはこの傾向が特に顕著である。屋久島で3月2-5日に掃立てた蚕を用い、4-5令期を硬葉(12月-1月発芽)で飼育した区と軟葉(2月下旬-3月上旬発芽)て飼育した区とに分け、15℃比重1.075と46℃比重1.075との塩酸液による浸酸成績を比較した処、軟葉区の孵化が悪く、塩酸については、早春採りに限らず15℃比重1.075によるのが安全であった(6,12)。
b 浸酸前冷蔵
浸漬を延期したい場合には、産卵後25℃で20時間目に5℃に冷蔵する。冷蔵可能の期間は最大限1週間で、なるべく短期間にとどめる。冷蔵車から出した後20℃-25℃ぐらいの温度の所に約2時間おいてから浸酸する。冷蔵中に卵が若干着色してくるが差支えない。
また、産卵直後から15℃で保護しておけば、浸酸適期を産卵後5曰ぐらいまでおくらせることができるから(第122表)、2、3日分の産卵をまとめて浸酸することができると云われているが(1149)、この場合には、上記冷蔵卵とは違い、産卵の翌日の浸酸は不可で、3日以後5日までの間に浸酸する。但し、渡辺の用いた日110号は非常に浸酸し易い品種で、現在の品種に比べると適期の巾が広いから、これをそのまま現行品種に適用することはやや危険である。
産卵後20時間目の卵には漿膜が未だ完成せず、乾燥に弱いから、冷蔵中の湿度に注意し、75%以下にならないように管理する。
c 脱酸、乾燥
浸酸の終った蚕種は直ちに水洗して、外部に付着している塩酸を除く。水温は20℃付近が最もよく、7
2℃以下の水で洗うと白ハゼ卵が出るが(673)、水温15℃-30℃の範囲では孵化に差がないと云う(1140)。苛性ソーダの1-2%液に浸して脱酸する方法もあるが、渡辺(1140)によれば、1%液に12分間以上、2%液に10分間以上浸したものは孵化が悪かった。9分間以下では被害がなかった。これは水温によっても影響があると考えられるが、記載されていない。
塩酸浸漬時間の不十分な蚕種は、脱酸せずにおいた方が孵化歩合が高かったと云う成績があって(1140)、孵化に関する限りは、脱酸は必らずしも厳格に行なう必要がないようであるが、脱酸不十分なものはその後の取扱い上不都合なばかりでなく、脱酸不十分な蚕種から孵化した蚕には多角体病の発生が多かったと云う報告もあるから(ⅩⅠ2)、舐めてみて酸味を感じなくなるまで水洗する。
脱水に遠心器を使用する場合の遠心力の影響は、卵令と廻転半径およひ廻転数によって相違があるが、松村ら(513,518)によれば、産卵後24時間の卵は廻転半径15cm、1分間3、000回、30分間ぐらいまでの廻転に耐えると云うから、耐えられる遠心力の限度は約1、500g、30分間と考えられる。遠心力(F)の強さは廻転半径および廻転数によって異なり、次式によって求められる。
F=s2 r/8,9500 (g)
rは廻転半径(cm)
sは1分間の廻転数
水洗後の乾燥が手間取ると孵化が悪くなる(578,580,732)。しかし、台紙着きの浸酸種を人工温度で乾かす場合に、卵面の濡れている間は28℃-29℃の高温の下でも卵に害はないが、卵面が乾いてから台紙の乾き切るまでをこのような高温下におくと、合紙の生乾きのときに25℃以下の温度に移したものよりも孵化が悪いと云う。これは、卵面の濡れている間は気化熱をとられるために、卵の温度が室温より低いが、卵面が乾くと卵が直接に高温を感じるためであろうとされている。
大櫛ら(732)が乾燥に要する時間が10時間以内ならば、その長短は孵化成績に大きな影響はないと云っているのは、台紙着きのものを1枚ずつにした場合のことで、濡れた台紙を積み重ねたり、濡れたバラ種を堆積しておくと、呼吸障害および蒸れのために被害がある。渡辺(1140)は、12時間ぐらい台紙を湿潤状態に保ち、以後8時間(計20時間)で乾かしたものは実用上差支えない孵化歩合を示したと云っているが、台紙着き蚕種を浸酸後2時間以内に乾かしたものと、6時間以内に乾かしたものとの浸酸後18時間目の胚の発育状態を比較すると後者がおくれていた(933)。胚の発育がおくれると、浸酸後の冷蔵成績に影響するおそれがある(Ⅷ2Cb)。
水野(578,580)は、水洗後の蚕種(台紙着き)を乾燥室内においた場合と同温度で湿度飽和状態の室においた場合との成績の比較から、乾燥のおくれた場合に孵化が不良なのは、水の蒸発により潜熱を奪われて卵が低温になることよりも、卵面が水に覆われて呼吸障害を来たすことに原因があるものと考えている。
水洗後、蚕種を長時間水に浸して置くと孵化か悪くなるが、この影響は水温によって相違する。滝沢・久保(1006)によると、水温20℃では1日で孵化歩合が80%ぐらいに低下し、その後4日で孵化歩合は0になったが、水温10℃の場合には9日で80%ぐらいになり、0になるのにはその後7日を要した。
d 比重1.10の塩酸液による加温浸酸
加温即時浸酸を15℃比重1.10の塩酸液を用いて行なうと、不受精卵がよく潰れて除去し易く、また塩酸液を冷蔵浸酸用と兼用できて無駄が言けると云う考え方がある(234,482,582-584,833)。
水田が長×光(日本種)および新玉(支那種)について産卵後18時間目、42時間目および66時間目に行なった15℃比重1.10の塩酸液による浸酸試験によれば、18時間目の液温40℃-42℃、5-7分間処理が適当で、孵化歩合は15℃比重1.075、46℃、5分間の浸酸(ただし産卵後18時間目浸酸)に劣らなかった。関(833)によれば、産卵翌日の午前11時に浸酸を行なう場合には100゚F(37 8℃)、6分間、午後4時に行なう場合には105゚F(40.6℃)6分間の浸漬がよかった(支107号)。沓掛(482)は日115号母体および支108号母体の交雑種について試験を行なったが、100゚F(37.8℃)では6-15
分間、110゚F(43.3℃)では6分間の浸漬がよかった。
これらの方法は実用的には殆ど用いられていない。
B 常温即時浸酸
塩酸液の濃度を高め、浸漬時間を長くすれば、液温は低くても浸酸効果をあが得ることは第130表によって明かである。無加温浸酸または室温浸酸と呼ばれている方法はこの原理によるものであるが、温度が低く過ぎると非常に長時間の浸漬が必要であり、従って、温度が不定であると孵化成績も不定であるから、実用上は25℃付近の液温を保つ必要がある。この意味で、無加温あるいは室温と云うよりは、常温浸酸と呼ぶのが適当と考えられる。
常温法の利点は、加温法に比べて、浸酸適期および有効浸酸時間の巾が広く液温の変動による被害が少ないことである。このため、日122号のように、加温法によって潰れ卵の生じ易い品種もよく孵化する(第132表)(125,447)。不受精卵の潰れもよい。
| 産卵後の時間 (時間) |
総孵化歩合(%) | 実用孵化歩合(%) | ||
| 加温浸酸 | 常温浸酸 | 加温浸酸 | 常温浸酸 | |
| 5 10 15 20 25 30 35 40 45 50 |
38 55 82 93 92 94 34 8 0 1 |
93 85 89 94 94 94 91 86 84 77 |
36 40 61 88 78 72 22 3 0 0 |
88 81 88 89 78 76 54 59 56 52 |
広部ら(206-208)は、常温法は、塩酸濃度を適当に選べば、産卵後3時間から80時間ぐらいまでの広範囲の卵令に亘って有効であり、日122号の浸酸にも適することを認め、河野・小川(455)は支124号×日124号を用い、産卵の不斉な場合の模型として、卵令10時間の卵10%、15時間の卵30%、20時間の卵50%、25時間の卵10%の割令の混合蚕種に常温浸酸を施し、孵化歩合98.8%、最多2日孵化歩合97.0%を得たと報告している。但し25時令を過ぎると孵化のおくれが目立ってくる(450)。
自然に産んだ卵の場合には、おそく産んだ、卵令の若いものほど卵が小さく(Ⅳ1Eb)、小形の卵は浸酸刺戟に弱い(Ⅷ1A)。人工的に卵令を混合する場合には、注意しないと、この卵の大小の関係を無視して、卵令だけの混合卵を作り、自然の場合とは著しく異なった材料で試験をするおそれがあるが、宮沢(572)によれは、蛹に大小があって蚕卵に大小を生じた場合にも常温浸酸がよいと云う。
常温浸酸の標準浸漬時間は第133表の通りである。
| 浸酸時間(時間) (産卵後の時間) |
液温別浸漬時間(分) | ||
| 24℃ | 27℃ | 29℃ | |
| 10 15 20 25 |
60-70 60-80 60-100 60-100 |
40-70 60-80 60-80 60-80 |
40 40-50 40-50 40-50 |
C 不越年種および即時浸酸種の冷蔵
a 不越年種
不越年蚕種を冷蔵した場合に、90%以上の孵化歩合の得られる有効冷蔵日数の限度は第134表の通りとされている。
| 冷蔵時期 | 有効冷蔵期間(日) | |||||
| 胚の状態 | 産卵後の時間 | -2.5℃ | 0℃ | 2.5℃ | 5℃ | 7.5℃ |
| 丙A* (Stage14) 丁A (Stage16) 丁B (Stage17) -戊A (Stage18) -戊B (Stage18) 戊B (Stage19) 戊C (Stage20) 己A (Stage21) 己C (Stage26) |
3日目 6時 〃 12時 〃 18時 4日目 0時 〃 6時 〃 12時 〃 18時 5日目 0時 9日目 6時 |
10 10 5 5 5 ○** 5 5 5 |
15 20 20 20 15 10 10 5 5 |
30 30 30 20 20 15 5 10 10 |
20 20 30 30 20 15 5 10 10 |
5 10 20 20 15 10 5 15 15 |
但し、今日では不越年蚕種の試験が殆ど行なわれていないので、これが現行蚕品種を不越年にした場合に、そのまま適用できるか否かは不明である。荒木ら(51)も75゚F(23.9℃)、45時間目(第134表の丁Bの時期に該当)に40゚F(4.4℃)に冷蔵するのがよいとしている(期間は夏蚕期30日以内、秋蚕期20日以内)。
水野は、第134表の発育段階別有効冷蔵期間を連結すれば、越年蚕種の場合と同様に(Ⅶ1Dc)、有効冷蔵期間を延長することができるとしている。1例を示せば、
丙A 2.5℃30 日+丁B 5℃20日=50日
の冷蔵によっても92.6%の孵化歩合を得たと云う。但し、丙Aで2.5℃に30日間冷蔵した後、一旦出庫して75゚F(23.9℃)で12時間保護し、胚を丁Bまで進めたものを5℃に20日間再冷蔵したのである。
不越年種の冷蔵については、なおⅧ4参照。
b 加湿即時浸酸種
加湿即時浸酸種の冷蔵法として普通に用いられるのは、
1)浸酸後 25℃、40時間目に2.5℃または5℃に冷蔵
2)浸酸後 25℃、18時間目に2.5℃に冷蔵
する方法である。
1)の冷蔵時期は、不越年種の冷蔵抵抗力の強い時期(丁B)にほぼ該当するが、即時浸酸種の胚の発育は浸酸の条件およびその後の取扱いによって異なるから、浸酸後の時間だけに頼ることは危険で、種々な条件を考慮する必要がある。三浦(547)は浸酸後75゚F(23.9℃)、約36時間目(6時間以内に風乾したもの)がこの時期に当ると云っている。胚を調べて冷蔵時期の参考にする場合には、付属肢が生じているか否かをみればよいから、越年種の場合よりは簡単で、生体解剖で十分間に合う。
胚の発育が丁Bを過きている場合には、2.5℃よりも5℃に冷蔵する方がよい。
実用上の冷蔵期間は20日以内と考えるのが安全である。
2)の方法は普通にダルマ形期冷蔵と云われているが、真の適期はダルマ形期(Stage5
)ではない。ダルマ形期に冷蔵すると畸形が発生し易く(128)、胚を調べて冷蔵してみると、ダルマ形期を過ぎてからの冷蔵の方が成績がよい(641)。
加温即時浸酸種がダルマ形期になるのは、普通、浸酸後15時間目頃で、浸酸が適当に行なわれ、その後の管理が適正であれば、浸酸後25℃、18時間の胚はダルマ形期を過きて、Stage6
になっている(933)。胚の発育は浸酸およびその後の取扱いの条件によって異なる。冷蔵成績の良否は冷蔵法そのものよりも、冷蔵時期の蚕種の状態によってきまると云ってよい。普通には、浸酸後18時間以内よりも18-20時間で冷蔵するのが安全であろう。
ダルマ形期直後が即時浸酸種の冷蔵適期であることは、小針・室賀(397)によって提唱されていたが、長野県蚕業試験場における試験成績によって(296,917)注目をひき、昭和32年度の地方蚕業試験場協力試験に取上げられて以来、広く用いられるようになった。
この協力試験においては、即時浸酸後、25℃、40時間目に2.5℃ (実際には各試験場によって一定せず、2.2℃-5.5℃の間で行なわれた)に冷蔵する区、同じく18時間目(胚子がダルマ形を呈する時期)に2.5℃
(実際には2.2℃-5.5℃)に冷蔵する区、および48時間目に5℃に冷蔵する冷浸区の比較が行なわれた。試験設計に“胚子がダルマ形を呈する時期”と明記されていたために、場所によっては正しいダルマ形期(冷蔵に不適当な時期)にわざわざ冷蔵したかも知れないこと、および冷蔵温度が、場所によっては、5.5℃と云うような高い場合もあったことに問題があるが、日122号×支115号、日124号×支124号およびその他の交雑種を用いての比較の結果として、次ぎのように結論されている。
孵化歩合 即浸種の40時間目区と18時間目区との間には、冷蔵35日までは差がなかったが、それ以後になると40時間目区が劣った。18時間目区と冷浸区との比較では、冷蔵期間30日以上になると後者が勝った。
減蚕歩合 冷蔵期間26日以上になると40時間目区が18時間目区に劣り、冷蔵35日になると、その差が明瞭であった。18時間目区と冷浸区との間には差がなかった。
経過日数 減蚕歩合に差の認められる頃の冷蔵日数になると、40時間目区が長くなる傾向を示したが、18時間目区と冷浸区との間には差がなかった。
収繭量 冷蔵期間26日以上になると40時間目区が18時間目区に劣り、冷蔵35日以上では18時間目区が冷浸区に劣った。
全繭重 冷蔵30日以上においては40時間目区が明かに劣り、18時間目区も漸次冷浸区より軽くなった。
繭層重 冷蔵30日以上では40時間目区が明かに18時間目区に劣り、冷蔵26日以上では18時間目区が冷浸区に劣った。
繭層歩合 冷蔵40日以上では40時間日区が18時間目区に劣ったが、18時間目区と冷浸区との間には差がなかった。
以上の結果を通覧すると、冷蔵期間30日以上の場合においては冷蔵浸酸が勝り、20日以上30日までの冷蔵では18時間目冷蔵がよかった。しかし、冷蔵35日ぐらいまでは18時間目区と冷浸区との間に大差はなかった。
水田ら(588,589)も加湿即時酸種の長期冷蔵試験を行なったが、浸酸後25℃、15-40時間の範囲内で冷蔵する場合には、早期の冷蔵には2.5℃よりも5℃がよく、冷蔵期間10-40日の範囲内において、浸酸が15℃比重1.075、46℃、1-7分間浸漬の場合には、浸漬時間の短かいほど長期間の冷蔵に耐えた。これは、浸酸による不越年化の不十分な卵においては、完全に活性化するまでの期間だけ耐冷蔵期間が延びたためと考えられるが、その代り、このような蚕種は短期間の冷蔵では孵化が悪い。また、この場合には、活性化を補う必要上、冷蔵温度は2.5℃よりも5℃がよく、冷蔵時期も浸酸後20-40時間の範囲内では早い方がよかった。浸酸刺戟が十分な場合には、活性化の問題には関係がなく、活力を維持するのが主体であるから、冷蔵温度は5℃よりも2.5℃がよい。これらの関係は、冷蔵期間を上記より長くしても同様であった。
堤・清水(1088)は、現行品種(原種)について試験を行ない、即浸後18-20時間目に冷蔵(5
℃、30日)したものの飼育成績が、40時間目冷蔵のものに比べて明かによいことを報告している。
しかし、冷蔵成績は蚕品種により、または原種と交雑種とによって著しく相違する(第135、136表)。
| 試験場所 (蚕期) |
冷蔵 | 試験 番号 |
浸漬時間 (分) |
日129号 | 支129号 | ||
| 初発蟻まで の日数(日) |
初発3日孵 化歩合(%) |
初発蟻まで の日数(日) |
初発3日孵 化歩合(%) |
||||
| 新庄 (春) |
無冷蔵 | 1 | 3 5 7 |
9 9 9 |
90 91 92 |
9 9 9 |
94 94 93 |
| 2 | 3 5 7 |
9 9 9 |
91 92 91 |
9 9 9 |
95 94 94 |
||
| 冷蔵 40日 |
1 | 3 5 7 |
9 9 9 |
89 88 84 |
9 9 9 |
90 90 90 |
|
| 2 | 3 5 7 |
8 8 9 |
90 90 85 |
9 9 9 |
77 80 87 |
||
| 宮崎 (春) |
無冷蔵 | 1 | 3 5 7 |
10 10 10 |
91 94 91 |
10 10 10 |
89 84 86 |
| 2 | 3 5 7 |
10 10 10 |
94 92 92 |
10 10 10 |
89 85 86 |
||
| 冷蔵 40日 |
1 | 3 5 7 |
10 10 10 |
88 76 67 |
10 10 10 |
78 75 74 |
|
| 2 | 3 5 7 |
9 9 9 |
94 79 60 |
10 10 10 |
81 83 77 |
||
| 試験場所 (蚕期) |
冷蔵 | 試験 番号 |
浸漬時間 (分) |
日129号×支129号 | 支129号×日129号 | ||
| 初発蟻まで の日数(日) |
初発3日孵 化歩合(%) |
初発蟻まで の日数(日) |
初発3日孵 化歩合(%) |
||||
| 新庄 (春) |
無冷蔵 | 1 | 3 5 7 |
9 9 9 |
94 95 96 |
9 9 9 |
97 96 96 |
| 2 | 3 5 7 |
9 9 9 |
95 95 97 |
9 9 9 |
97 98 97 |
||
| 冷蔵 40日 |
1 | 3 5 7 |
8 8 8 |
95 95 95 |
9 9 9 |
97 96 96 |
|
| 2 | 3 5 7 |
8 8 8 |
95 95 92 |
8 8 8 |
92 94 95 |
||
| 宮崎 (春) |
無冷蔵 | 1 | 3 5 7 |
10 10 10 |
97 97 94 |
10 10 10 |
92 93 93 |
| 2 | 3 5 7 |
10 10 10 |
97 97 97 |
10 10 10 |
92 93 94 |
||
| 冷蔵 40日 |
1 | 3 5 7 |
9 9 9 |
96 88 74 |
10 10 10 |
90 93 90 |
|
| 2 | 3 5 7 |
9 9 9 |
97 92 84 |
9 9 9 |
84 89 91 |
||
即時浸酸種の長期冷蔵によって蚕作の不良になった場合を、長野県蚕業取締所(634)の調べた実例によって示すと次ぎの通りである。
冷蔵20-25日のものに、毛振いをしない蚕があり、眠起不斉で稚蚕期の減蚕が多かったが、不良蚕は3眠までに除去され、蚕作に大きな影響がなかった。冷蔵40日のものでは(支105号×日110号)、4農家合計222gの掃立てに対し、収繭量22貫160匁、対蟻量1g収繭量は99匁で、殆で全滅に等しかった。但し、冷蔵方法などには言及していない。
c 常温即時浸酸種
加温即時浸酸においては、高温で処理するため、胚の発育が一時的には無処理の越年種よりも却っておくれる。
産卵後25℃、20時間目(Stage4 )に加温浸酸を施すと、胚がダルマ形(Stage5
)になるまでに15時間(産卵後35時間)かかるが、無浸酸の越年種は産卵後30時間余りで同じ段階に達する。
常温浸酸を施した蚕種においては、胚の発育が本来の不越年種におけるよりはおそいが、加温浸酸種のようなおくれはなく、産卵後30時間でダルマ形期に達するから、冷蔵時期を加温法の場合よりも早める必要がある。即ち、加温法においては、産卵後20時間目に浸酸し、その後18-20時間、計38-40時間目が所謂ダルマ形期冷蔵(実際にはダルマ形を過ぎている)の適期であるのに対し、常温法においては、産卵後24℃、15時間日から15℃比重1.110、液温25℃、75分間の浸漬を行ない、その後25℃で保護した場合には、浸酸後18時間45分、産卵後通計35時間目が冷蔵適期であって、蚕品種によって程度は異なるが、産卵後40時間目に冷蔵すると、35時間目の冷蔵よりも成績が劣った(452,453)。
3 冷蔵浸酸
A 冷蔵浸酸の基本問題
冷蔵浸酸は、蚕種を産卵後2日目前後に5℃に冷蔵し20日以上たってから出庫、浸酸するのが普通の方法であるが、この冷蔵当時の蚕種に浸酸を施こすと孵化は若干不斉であるが孵化能力は十分にある(第122表)。従って、冷蔵浸酸における冷蔵は、越年性になった卵の活性化を進める人工越冬よりも、孵化能力のある蚕卵の冷蔵であるとする考え方がある(775)。人工越冬浸酸法と呼ばずに、冷蔵浸酸法と呼ばれるようになったのはこの理由によるものと思われるが(580)、産卵後2日間も高温に保護し、越年化の進み初めた蚕種を5℃に冷蔵すると、越年化の進行は直ちに停止する訳ではなく、弱い浸酸を施して調べると、寧ろ冷蔵後20日間ぐらいは孵化歩合が次第に低下することがわかる(第137表)。
| 冷蔵期間 | 浸 酸 | 総孵化歩合 (%) |
最多2日孵化 歩合(%) |
| 16時間 | 比重1.110、48℃、5分間 比重1.120、常温、1時間 〃 〃 〃 4時間 〃 〃 〃 14時間 |
0 87 42 0 |
0 67 38 0 |
| 6日間 | 比重1.110、48℃、5分間 比重1.120、常温、1時間 〃 〃 〃 4時間 〃 〃 〃 14時間 |
19 84 80 9 |
14 59 66 8 |
| 10日間 | 比重1.110、48℃、5分間 比重1.120、常温、1時間 〃 〃 〃 4時間 〃 〃 〃 14時間 |
5 35 87 70 |
4 21 61 51 |
| 15日間 | 比重1.110、48℃、5分間 比重1.120、常温、1時間 〃 〃 〃 4時間 〃 〃 〃 14時間 |
23 3 61 76 |
18 3 41 60 |
| 20日間 | 比重1.110、48℃、5分間 比重1.120、常温、1時間 〃 〃 〃 4時間 〃 〃 〃 14時間 |
16 2 66 84 |
13 2 42 60 |
この表の常温1時間浸漬をみると、冷蔵16時間後には87%も孵化したものが、冷蔵6日、10日と次第に孵化歩合の低下を示し、冷蔵20日間では2%に下がっているから、孵化歩合からみる限り、少なくとも冷蔵20日までは越年化の進行が停止していないことが明かである。同様な現象は休眠に伴なうグリコーゲンの減少(Ⅴ2B)にも認められる。越年性卵を産卵後2日目に5℃に冷蔵すると、高温保護のものに比べてクリコーゲンの減少は明かに低下するが、減少は20日以後まで続き、30日ぐらいで漸く最低値に達する。
これらのことからみると、第137表において、加温浸酸および常温14時間浸漬のような強い処理を施こすと、冷蔵16時間では殆ど0であった孵化歩合が、冷蔵6日で加温19%、常温14時間9%に上り、冷蔵20日で前者16%、後者84%になっているのは蚕種の活性化によるものではなく、抵抗力の増大によるものと考えなければならない。冷蔵中に蚕種の浸酸抵抗力の増加することは、若上げした日122号の卵が46℃比重1.075の即時浸酸によっては潰れ易いにも拘らす、冷蔵浸酸によっては潰れにくいこと(1072)からも明かである。
冷蔵浸酸を冷蔵期間の長短によって区別し、冷蔵期間40日未満のものを短期冷蔵浸酸、40日以上のものを普通または長期冷蔵浸酸と呼んでいるのは、一見、極めて便宜的な区別のように思われる。しかし、上記のことから考えると、これは冷蔵浸酸の本質にふれた重要な区別である。
短期冷蔵浸酸は越年化の進行中、または停止する以前の蚕種の人工孵化であり、長期冷蔵浸酸は不完全人工越冬卵の活性化を浸酸処理によって補なう方法であることを根底として問題を考えて行かなければならない。
一般に、冷蔵期聞が短かいと活性化処理期間が不十分な訳であるから、冷蔵期間の長い場合に比べて早目(越年化の進まないうち)に冷蔵するのがよいと考えられている。水野(580)はこの点について次ぎのように述べている。
冷蔵浸酸における冷蔵処理は最も有効な人工越冬を行なうことである。人工越冬としては、保護温度が75゚F(23.9℃)内外の場合には、産卵後3日乃至4日目(産卵当日を第1日とする)に5℃または7.5℃に冷蔵するのがよいが、冷蔵適期は日数ではなく、胚の発育程度によるのであるから、初期発育のおくれる一化性品種の冷蔵適期は4日目早朝を中心とし、二化性品種は3日目夕を中心にするのがよい。従って、冷蔵浸酸にもこの時期の冷蔵が有効な訳である。しかし、冷蔵20日以内の短期冷蔵浸酸においては、上に述べたこととは若干喰違うが、即時浸酸の有効な時期に近付ける意味で、幾分早目に冷蔵する方が成績がよい。
このような考えに基づいて水野が実際に示している標準は、産卵後の保護温度75゚F(23.9℃)の場合に、
短期冷蔵浸酸 3日目夕(小豆色)に40-45゚F(4.4℃-7.2℃)
冷蔵期間40-50日の場合 3日夕乃至4日目早朝(小豆色)に40-45゚F
であって、短期の冷蔵時期を40-50日間冷蔵のものに比べて特別に早めてはおらず、40-50日間冷蔵の場合に適期の巾があと(4日目早朝)へ延びているだけである。
産卵後の保護温度と冷蔵適期との関係についての渡辺の試験結果は既に述べたが(Ⅷ1A)、渡辺は冷蔵期間と冷蔵適期との関係についても試験を行ない、冷蔵期間の短かい場合ほど適期の巾の狭いことを述べている。即ち、日110号を産卵後、温度25℃、湿度75%で保護し、5℃に冷蔵した場合の適期(47.8℃比重1.10の塩酸液で5分間の浸漬により90%以上の孵化の得られた)の巾は
冷蔵期間 10日の場合 産卵後 2日-2日半
〃 15日 〃 〃 2日-3日
〃 20日 〃 〃 1日半-4日
〃 30日 〃 〃 1日半-10日
〃 40日 〃 〃 1日-30日
であった。このように適期の巾が広ければ、冷蔵浸酸には何の苦心もいらないぐらいで、これを現在の蚕品種一般に適用することはできないが、一般的な傾向としては重要な結果であって、長期冷蔵の冷蔵適期が短期冷蔵の適期よりも巾が広いばかりでなく、長期の巾は短期の巾よりも早い方へも拡がっている点で注目される。
足立(15)の日107号×支101号、日107号×(支4号×支101号)および日107号についての試験成績においても、40、45および50時間目の冷蔵結果を比較すると、冷蔵時期の早いものほど短期冷蔵浸酸の成績が悪い。但し、この場合の浸酸は塩酸濃度20%、液温48℃、5分間浸酸であった。
このように、短期冷蔵浸酸の冷蔵時期は早目にするのがよいと云う考えと必らずしも一致しない結果の多い原因は何であろうか。
この点を明かにするためには、冷蔵時期と共に塩酸刺戟の強さをも変化させた試験結果を検討する必要がある、第138表およひ第139表はこの問題に関して興味のある成績である。
| 塩酸濃度 (%) |
冷蔵時期 (時間目) |
総孵化歩合平均(%) | 実用的孵化斉否率(%) | ||||
| 2.5℃ | 5℃ | 7.5℃ | 2.5℃ | 5℃ | 7.5℃ | ||
| 18 | 30 40 50 60 |
66 74 89 87 |
83 88 87 87 |
78 77 74 73 |
89 88 86 81 |
81 84 86 81 |
68 66 64 63 |
| 20 | 30 40 50 60 |
44 61 87 91 |
82 95 95 94 |
90 89 92 90 |
87 88 85 82 |
88 88 87 86 |
74 70 71 68 |
| 22 | 30 40 50 60 |
34 54 84 83 |
86 95 95 95 |
93 92 93 90 |
89 89 88 86 |
90 90 91 90 |
82 84 83 83 |
| 塩酸濃度 (%) |
冷蔵時期 (時間目) |
総孵化歩合平均(%) | 実用的孵化斉否率(%) | ||||
| 2.5℃ | 5℃ | 7.5℃ | 2.5℃ | 5℃ | 7.5℃ | ||
| 16 | 35 40 45 50 55 60 |
63 70 65 50 30 21 |
83 87 85 84 71 65 |
86 84 81 72 64 53 |
59 51 40 33 25 17 |
41 36 36 39 35 24 |
28 26 32 34 28 24 |
| 18 | 35 40 45 50 55 60 |
67 79 86 84 78 67 |
89 94 93 93 88 78 |
94 90 87 83 76 66 |
78 73 69 70 50 40 |
62 61 66 58 46 29 |
48 44 49 37 32 25 |
| 20 | 35 40 45 50 55 60 |
67 85 91 92 90 90 |
90 95 95 95 92 92 |
95 92 93 94 89 88 |
81 82 84 85 68 72 |
85 77 78 75 59 61 |
66 63 64 52 44 47 |
| 22 | 35 40 45 50 55 60 |
76 84 92 92 93 93 |
91 96 95 96 94 91 |
96 92 94 95 94 93 |
83 84 90 94 79 77 |
90 90 91 93 91 61 |
79 88 74 85 74 60 |
室賀(611)はこの結果から、二化性品種の冷蔵浸酸において、産卵後を24℃に保護し、冷蔵日数30日間の場合には、産卵後40-60時間目に5℃に冷蔵し、出庫後塩酸液の濃度22%、液温48℃、浸漬5分の浸酸を行なうのがよく、産卵後25℃保護、冷蔵期間40日の場合には産卵後40-55時間目に5℃に冷蔵し、上と同様の浸酸をするのがよかったと云い、2.5℃に冷蔵すると、孵化歩合は低いが孵化は5℃と同様に斉一なこと、7.5℃冷蔵においては、孵化歩合は低くないが孵化不斉の傾向のあること、また、冷蔵時期のおそいものは早いものに比べて、孵化歩合は高いが孵化比不斉の傾向のあることなどを指摘している。
冷蔵浸漬の場合に7.5℃冷蔵が5℃冷蔵より劣る理由について、室賀(622)は活性化を促進する温度の順位が7.5℃、5℃、10℃、2.5℃であるにも拘らず(第96表)、冷蔵浸酸においては5℃冷蔵が7.5℃冷蔵に勝っているのは、低温による活性化と塩酸による活性化とが本質的に異なり、低温によって活性化し易くなった卵が必らずしも塩酸刺戟によって活性化し易い卵ではないためであるとしている。塩酸による活性化は主として卵原形質内に存在する抑制質が破壊されるためであり、低温による活性化は卵原形質並びに胚内の抑制質が消費されるためであるから、7.5℃においては5℃におけるよりも消費が多く、残存する抑制質は少ない筈であるが、7.5℃においては同時に胚の発育も進むため、胚内に移行する抑制質量が多く、体内に移行したものは塩酸によっては破壊し難くなると云うのである。
室賀のこの仮定は現在のところ証明することができないから(Ⅴ2A)、抑制質の移行その他の問題には触れ得ないが、7.5℃ばかりでなく、5℃においても冷蔵中に胚の発育が進み(15,230)、越年化の進んでいることは前述の通りである。従って、5℃に冷蔵した卵の卵内状態が、7.5℃に冷蔵した卵のものとは全然相違していると云う前提で考えることには無理がある。
冷蔵30日間の場合(第138表)において注目されるのは、塩酸濃度の如何にかかわらず、2.5℃冷蔵は、冷蔵時期のおそいほど孵化がよいこと、および5℃ならびに7.5℃冷蔵とは違い、塩酸濃度を高めても孵化歩合が目立ってよくならず、22%の場合には却って孵化歩合の幾分低下するようにさえみえることである、これは抑制質の破壊だけによっては説明が困難で、室賀は孵化しなかった卵の状態を記載していないが、第137表の場合と同様に、浸酸に対する抵抗力と塩酸刺戟の効果との釣合ったときの孵化歩合が最も高いことを示しているものに外ならないと思われる。
7.5℃冷蔵においては、冷蔵中の抵抗力の増加も越年化の進み方も2.5℃に比べて速いから、この釣合いは、2.5℃の場合に比べて、冷蔵時期の早い、塩酸刺戟の強い条件の下で成立する筈であるが、実際に、孵化歩合の高いのはこのような条件のときである。
これに対して5℃冷蔵においては、これら両極端の中間に当る比較的範囲の広い条件の下で釣合いが成立するために、孵化が最もよかったと考えられるのである。
2.5℃冷蔵において、孵比歩合は低いが孵化が斉一なのは、5℃または7.5℃に比べて抵抗力の増加はおそいが、越年化の進行が抑えられていることを示すものであろう。
このような考え方で冷蔵40日の場合(第139表)をみると、冷蔵30日の場合とは材料が相違すること、産卵後の保護温度が1℃高かったこと、不越年化が進んでいる筈であること、浸酸抵抗力が増加して、2.5℃冷蔵のものの孵化の様相が30日間冷蔵の場合の5℃冷蔵に近付いたことなどの相違点があって、上と同一には論じ得ないか、塩酸濃度の特別に低くかった16%の場合を除けば、2.5℃冷蔵には冷蔵時期の早いものに孵化の悪い傾向が残っており、7.5℃冷蔵には冷蔵の早いものの孵化歩合が高く、5℃冷蔵はこれらの中間で、釣合いの成立する条件の範囲の広いことには変りがない。
冷蔵期間が40日になると、2.5℃においても、塩酸度度20%以下では55時間目以後の冷蔵の孵化が悪いこと、30日冷蔵に比べて孵化の不斉なことなどをみると、2.5℃においてもまた浸酸抵抗力の増加と共に越年化の若干進むことがうかがわれ、2.5℃、5
℃、 7.5℃の冷蔵効果の間に、室賀の考えているような本質的な違いはないもののように思われる。
大櫛ら(734)は、産卵後27℃、40時間目に4.5℃に冷蔵すると、10-40日間の冷蔵浸酸によって90%以上の孵化歩合を得たと云っているが、10日間冷蔵のものは塩酸濃度20%、115゚F(46.1℃)、7-10分間浸漬であるから、液温を下げて釣合いをはかっていることがわかる。
仲野(657)は冷蔵期間15、20、25、30日の冷蔵浸酸を試験したが、その結果は浸酸抵抗力と塩酸刺戟の効果との釣合いが大切で、単に冷蔵時期を早めるとか塩酸刺戟を強めるだけでは短期冷蔵浸酸の孵化成績が改善されないことをよく示している。例えば冷蔵15日の場合、産卵後の保護温度20℃のときには、冷蔵温度が2.5℃ならば産卵後48-60時間目、5℃および7.5℃ならば36時間目にそれぞれ冷蔵し、保護温度24℃のときには、2.5℃および5℃ならば48時間目、7.5℃からは24時間目に冷蔵するのが最もよかった。塩酸刺戟(比重1.10、48℃)も弱めて、出庫後5時間目に3分間浸漬するのがよかった。
この仲野の2.5℃、15日間冷蔵の場合は即浸前冷蔵の変形とも考えるべきものである、冷蔵期間が短かく、活性化の余俗のない場合には、活性化の促進よりも越年化の進行を抑えることに重点をおく方法も考えられるのであって、前記の大和田(775)の考えはこの意味の冷蔵浸酸である。
一般に、冷蔵浸酸の冷蔵適期は卵色が小豆色の時期であるといわれている。これは三浦の研究結果によるのてあるが、三浦(547)は無条件に小豆色の時期がよいといっているのではない。日107号を産卵後76.5゚F
(24.7℃)に保護した場合の卵色および、これを40゚F(4.4℃)に冷蔵したときの耐久力は
産卵当時 黄色 3日-2週間
産卵後32時間内外 淡赤色 1カ月内外
〃 40 〃 赤色 2カ月以上
〃 48 〃 小豆色 ┐
〃 56 〃 殆ど固有色 ├数ヶ月間
〃 80 〃 固有色 ┘
で、これを浸酸すると、黄色および淡赤色で冷蔵したものは、方法がよければ殆ど全部一斉に孵化させることができるが(1日に約80%)、赤色期に冷蔵すると孵比がやや不斉で、出盛りが2日に亘り、小豆色冷蔵では更に不斉であった。しかし、冷蔵期間が30-40日になると小豆色冷蔵でも出盛りを2日以内に短縮することができる。従って、短期冷蔵(10日以内)のものは冷蔵を早目にするのがよいが、これに対する浸酸は、塩酸濃度は同じ15℃比重1.10でも、冷蔵20日以内のものには液温を低くし、浸漬時間を長くしている。卵色小豆色のときに40℃に冷蔵し、出庫後3時間以内に15℃比重1.10の塩酸液を用いて浸酸する場合の標準は
| 無冷蔵(小豆色) | 110゚F(43.2℃)、10-12分、または 115゚F(46.1℃)、6分 |
| 冷蔵10日(出庫当時の卵色は小豆色よりやや濃色) | 115゚F、10分、または 117゚F(47.2℃)、5分 |
| 冷蔵20日(固有色に近い) | 115゚F、8分、または117゚F、4-5分 |
| 冷蔵30日(固有色や、色素やや点集するものあり) | 118゚F(47.8℃)、4-5分 |
| 冷蔵40日 | 118゚F、5分 |
| 冷蔵50-60日(固有色、色素の点集漸次大となる) | 118゚F、6分 |
であって、極く短期の冷蔵浸酸の場合に、仲野が、液温を変えずに、出庫後の時間を長くし、浸漬時間を短かくして釣合いを求めたのに対し、三浦は液温を下げ、浸酸時間を長くして、一見相反するようではあるが、矢張り釣合いの条件を求めているのである。三浦の考え方は、常温浸酸によって即時浸酸の潰れ卵を少なくすると同じ発想である。
短期冷蔵については、孵化しにくい卵であるから十分強い浸酸処理が必要であるという意見と、ゆるく柔かに処理するのがよいという意見とがあって、相反する考え方のように思われがちであるが、これは釣合いの条件を何処に求めるかという方法の違いに過ぎないのであるから、形だけにとらわれることなく、その根本に共通する考え方を理解する必要がある。なお、室賀が第138、139表の試験に用いた22%の塩酸液は、47.8℃比重1.10の塩酸液と殆ど等しいか、むしろ低目の濃度であって、特別に強い塩酸刺戟を与えている訳ではない。
このように考えてくると、冷蔵後、なお越年化の進行している期間内における冷蔵浸酸(短期冷蔵浸酸)は、現在の処、積極的に不越年化を図る手段がないために、越年化を進めず、卵を潰さないという消極的な枠の中で釣合いを求めているのに過ぎないから、その成績は蚕種の生理状態によって殆ど決定されてしまう。冷蔵適期、冷蔵適温、有効浸漬時間なとの巾の狭いのはこのためである。
鎌倉(303)は、産卵後36-40時間目に-10℃~-17℃に48時間接触(第一次冷蔵)させた後、7℃に冷蔵(第二次冷蔵)すると、第一次、第二次合計20日以内の冷蔵の場合においても、従来の冷蔵浸酸法に比べて孵化がよいと報告しているが、従来の方法として用いたのは、産卵後48時間より早い時期に5℃に冷蔵し、比重1.100、液温48℃、5分間の浸酸であり、新らしい方法では、同じ時期から冷蔵したものに比重1.126、液温28℃、90分間の浸酸を施こしているのであって、後者は常温浸酸に近い処理であるから、従来の方法に潰れ卵が多く、新らしい方法より劣るのは当然で、これまで述べてきた釣合いの考え方からみれば、対照の取り方が適当でないように思われる。
しかし、この方法による成績には注目される点が一つある。第二次冷蔵を7℃にすると5℃よりも孵化成績のよいのは、発育が進んで抵抗力が増大するためと考えられないこともないが、総孵化歩合ばかりでなく、最多2日孵化歩合も7℃冷蔵において高いのをみると、第138表の成績と比べて、あるいは第一次冷蔵には越年化を不越年化の方向に転換させるか、または越年化の進行を停止させる作用があり、このために第二次冷蔵を7℃にしても孵化が不斉にならないのではないかということである。この点を論議する証拠は示されていないが、後に述べること(Ⅷ5A)とも関連して興味がある。なお鎌倉は、第二次冷蔵を長くすれば、短期冷蔵をそのまま、長期冷蔵に切り換えることができるといい、第二次冷蔵を7℃のまま63日間続けた例を示しているが、これを一般の品種にも適用し得るか否かについては検討の余地があるように思われる。
室賀(619)は、先きに述べた仮定から出発して、抑制質の胚子体内への移行を妨げれば冷蔵浸酸種の孵化成績がよくなるであろうと考え、産卵後25℃、48時間目に、水のしたたらない程度に濡らした蚕種をシャーレに満たし、その中央に試験用の蚕種を入れて蓋をした(濡卵)区、乾いた卵をシャーレに満たし、その中央に試験用蚕種を入れて蓋をした(乾卵)区、およびシャーレに入れたのみで蓋をしなかった対照区とを5℃に冷蔵し、20-40日間冷蔵した後に濃度22%、液温48℃、5分間の浸酸を施こし、孵化を比較した。それによると、濡卵区の孵化が最もよく、乾卵区がこれに次ぎ、対照区の成績はこれらに比べて明かに劣っていた。室賀はこの結果を、冷蔵中の呼吸障害によって、抑制質の胚子体内への移行が妨けられた結果、濡卵区の成績がよくなったものと考えている。但し、冷蔵中の湿度の影響でなかったか否かの吟味がないのて、未だ確実な結果とは云われない。
短期冷蔵浸酸種を、出庫、浸酸後25℃ に保護し、6-36時間目の間に35℃(湿度60-70%)に12-30時間接触させると孵化が斉一になると云われているが(476)、実用的な効果は得られなかったと云う報告もある(770,771)。但し、浸酸によって被害を受けた卵が高温接触によって回復する訳ではないから、効果の有無はこの点を考慮して判定しなければならない。この高温接触は蚕作に悪影響があると云われている。
冷蔵中の蚕種が浸酸によって十分孵化し得るようになったか否かを判定する方法として、足立(15)は漿膜に現われる黒点を目標にし、これが明瞭になったときに浸酸すれば孵化成績がよいと云っている。安喰(24)は漿膜色素粒の状態によって
1)色素粒発現期 微細色素粒が均一に現われた時期
2)色素粒成長期 色素粒が1)より成長し、2~3粒宛集合して大きさを増す時期(色素粒に大小の生じたときを前期、色素粒に大小が相半ばするときを中期、色素粒に大が多く、小が少ないときを後期とする)
3)色素粒凝集期 色素粒が2)よりも集合し、肉眼で観察される時期(前、中、後期に分ける)
の3時期を区別し、浸酸成績は成長期中期以後がよいとした。昭白、新光および昭白×新光を産卵後75゚F(23.9℃)、48時間目に5℃に冷蔵した場合には、冷蔵20-22日で発現期、28-29日で成長期、35-37日で凝集期になったと云う。
これらの方法が取上げている形態変化は、上に述べた蚕卵の浸酸抵抗力の指標であるのか、越年化の停止または不越年化への転換を意味するのかが不明であり、また形態的特徴そのものの発現条件も十分明かにされていないので、低温下における卵の発育に件なう平行現象であるのか、原因に結び付いた意味があるのかは不明であって、その普遍性も明かでない。
冷蔵した蚕種において、何日ぐらいで越年化の停止または不越年化への転換が起るかは、短期冷蔵浸酸と長期冷蔵浸酸との取扱い上重要な問題である。人工越冬の成績によると、産卵後48時間目に5℃に冷蔵した蚕種を、出庫後そのまま催青すると、品種にもよるが、冷蔵20日では殆ど孵化機能が認められず、30日では明かに孵化機能が現われている(Ⅷ6)。
グリコーゲンの減少の停止時期については先に述べたが、核分裂の消長も注目される。日124号6蛾、および支124号8蛾、の卵を産卵後25℃、40-50時間目に5℃に冷蔵して調べた結果によると(386)、25℃において前休眠期の細胞核分裂の終息するまでには3日かかるから、冷蔵当時には未だ分裂が行なわれている。冷蔵5日後からときどき出庫して、毎回1蛾につき10粒宛の卵を調べてみると、初めの間は、出庫後2時間では核分裂が認められるが、24時間後には認められなくなる状態が続いた。これは、冷蔵によって抑えられていた前休眠期核分裂の残りが出庫後1日以内に終了して、卵が休眠に入ることを示している。処が、支124号においては冷蔵15日、日124号においては冷蔵30日以後になると、この状態が変って、出庫後1日以上たっても核分裂が認められるようになる。これは、これらの卵が既に完全な越年性卵ではなくなり、後休眠期核分裂を開始したことを示している。但し、後休眠期核分裂が始まったとは云っても、完全な孵化機能を得た意味でないことは越年卵の活性化初期と同様である(Ⅵ2Bb)。
冷蔵中の蚕種の炭酸ガス排出量(25℃に24時間おいてから測定)を測定すると、最低値に達するのは冷蔵後20-30日目頃で、その後増加の傾向を現わす(872)。
これらの結果は越年性の転換が冷蔵後20-30日目頃に起こることを示しているが、冷蔵浸酸成績の安定し初めるのもこの頃である。
冷蔵浸酸の予定を中止して、蚕種を越年用に転用し得るのは冷蔵20日頃までであると云われているのも(523,580)、転換期が普通には20日以後であることを物語っているが、支124号のような品種ではこれより早いと考えられるから、注意しなければならない。
B 冷蔵浸酸の施行標準
a 冷蔵期間40日以上の場合
蚕種を、産卵後25℃に保護し、48-50時間目に5℃に冷蔵する。但し、産卵後の時間を20時を基点に数え、それ以前の産卵を用いる場合にはこれより、3時間ぐらい早目に冷蔵するのがよい。冷蔵期間50日以上を予定している場合には20時基点のままで差支えない。産卵中および産卵後の保護温度が違うと産卵後の時間だけでは冷蔵時期がきめられないから、積算温度によって取扱いをするのがよいと云う考えもあるが、保護温度が違えば冷蔵適期までの積算値も相違するので、その値だけに頼る訳には行かない。保護温度を一定にすることが総ての場合に大切である。積算温度は蚕品種によっても幾分異なる(1038a)。日欧固定種母体の蚕種を45日以上冷蔵する場合に、冷蔵時期が早過ぎると孵化はよいが蚕は弱いと云う報告があるが(360)、一般の品種においても、冷蔵期間60日以上の予定のものは、冷蔵時期を産卵後50-60時間におくらせる。冷蔵温度も、期間が40日以上になる場合には40日で2.5℃に下げる。
保護温度が30℃以上になっているような場合には、中間温度(15℃)に約2時間おいてから冷蔵するのが安全であるが、25℃に保護中のものは直接5℃に冷蔵しても、実用上、害はない。
冷蔵車の湿度は75-80%を目標とし、これ以下に下らないようにする。湿度の高い方が孵化がよい。
浸酸は、出庫後20-25℃の中間温度に2時間ぐらいおいてから行なう。塩酸は15℃比重1.10(約20%)、液温48℃
とし、もし48℃で比重を調整する場合には1.085とする。但し古い塩酸を混ぜない場合である(Ⅷ1Bc)。48℃比重1.10で浸酸を行なうことも多いが、この場合にもし潰れ卵が出るようであれば、濃度が高過ぎるのである。
浸漬時間は品種や冷蔵期間によっても異なるが、一般的に云えば
1化性品種 6-8分
2化性品種 5-7分
で、交雑種の浸漬時間は両親の中間でよいが、産卵後の時間がたっているために、即時浸酸種の場合よりは父の影響のかなり強く出ることがある。
現行品種についての実例を第140、141表に示す。冷蔵30日と40日との間で急に孵化歩合の上昇していることが認められる。
| 試験場所 (蚕期) |
冷蔵 日数 (日) |
試験 番号 |
浸漬 時間 (分) |
日129号 | 支129号 | ||||
| 初発蟻ま での日数 (日) |
初発3日 孵化歩合 (%) |
初発5日 孵化歩合 (%) |
初発蟻ま での日数 (日) |
初発3日 孵化歩合 (%) |
初発5日 孵化歩合 (%) |
||||
| 新庄 (春) |
30 | 1 | 5 7 9 |
10 10 10 |
93 92 94 |
94 93 95 |
11 11 11 |
88 92 92 |
93 93 93 |
| 2 | 5 7 9 |
10 10 10 |
97 98 95 |
97 98 95 |
11 11 11 |
87 86 80 |
89 88 83 |
||
| 40 | 1 | 5 7 9 |
10 10 10 |
93 93 95 |
94 94 95 |
10 10 10 |
78 85 96 |
95 94 97 |
|
| 2 | 5 7 9 |
10 10 10 |
96 95 96 |
96 96 96 |
11 11 11 |
91 94 89 |
92 95 90 |
||
| 宮崎 (春) |
30 | 1 | 5 7 9 |
11 11 11 |
95 96 96 |
97 97 97 |
12 12 12 |
65 73 71 |
79 83 80 |
| 2 | 5 7 9 |
11 11 11 |
95 96 97 |
98 98 98 |
12 12 12 |
57 63 69 |
70 74 78 |
||
| 40 | 1 | 5 7 9 |
11 11 11 |
94 97 97 |
97 98 97 |
12 12 12 |
86 88 87 |
91 92 91 |
|
| 2 | 5 7 9 |
11 11 11 |
95 97 97 |
97 98 98 |
12 12 12 |
88 90 90 |
93 94 93 |
||
| 試験場所 (蚕期) |
冷蔵 日数 (日) |
試験 番号 |
浸漬 時間 (分) |
日129号×支129号 | 支129号×日129号 | ||||
| 初発蟻ま での日数 (日) |
初発3日 孵化歩合 (%) |
初発5日 孵化歩合 (%) |
初発蟻ま での日数 (日) |
初発3日 孵化歩合 (%) |
初発5日 孵化歩合 (%) |
||||
| 新庄 (春) |
30 | 1 | 5 7 9 |
10 10 10 |
98 97 96 |
98 98 96 |
10 10 10 |
97 98 97 |
98 98 98 |
| 2 | 5 7 9 |
10 10 10 |
98 99 98 |
98 99 98 |
10 10 10 |
96 93 81 |
98 94 83 |
||
| 40 | 1 | 5 7 9 |
10 10 10 |
96 97 97 |
97 97 98 |
10 10 10 |
98 98 98 |
99 98 99 |
|
| 2 | 5 7 9 |
10 10 10 |
98 98 98 |
98 98 98 |
10 10 10 |
97 92 79 |
97 93 79 |
||
| 宮崎 (春) |
30 | 1 | 5 7 9 |
11 11 11 |
94 95 95 |
96 95 95 |
12 12 12 |
87 88 89 |
94 94 94 |
| 2 | 5 7 9 |
11 11 11 |
96 97 97 |
98 98 98 |
12 12 12 |
75 79 79 |
89 89 88 |
||
| 40 | 1 | 5 7 9 |
11 11 11 |
96 95 96 |
97 96 96 |
12 12 12 |
97 95 95 |
97 98 97 |
|
| 2 | 5 7 9 |
11 11 11 |
96 98 97 |
97 98 97 |
12 12 12 |
94 95 93 |
96 96 95 |
||
鎌倉(302)は、冷蔵期間45日以上の場合には、比重1.126または1.128の常温浸酸も実用性があると云っている。
上蔟後、産卵中および産卵後の温度に注意すること、集中産卵を図って卵令の巾を小さくすることなどは即時浸酸の場合以上に大切である。
b 冷蔵期間40日未満の場合
短期冷蔵浸酸の基本的な問題については既に詳述したが、それによってみても、また即時浸酸種の冷蔵成績から考えても、蚕種そのものだけを考えれば、冷蔵20日以内は即時浸酸冷蔵種を用いるのがよい。更に、第140、141表と第135、136表とを比較すれば、少なくとも冷蔵25日、品種によっては冷蔵30日でも即時浸酸冷蔵種の用いられることがわかる。しかし、即浸冷蔵種は、使用期限を25日から30日まで延長できたとしても、その先きがないのであるから、30日以後に孵化歩合が急に向上する冷蔵浸酸種との何れに比重をおくかは、技術面からばかりではなく、経営上の問題として考えなければならない。ここでは冷蔵25日以上の場合を対象にして述べる。
冷蔵期間30-40日の場合には、産卵後25℃、48時間目(20時を基点としてそれ以前の産卵を用いる場合には約3時間早める)を目標にして、蚕種を5℃に冷蔵する。冷蔵中の温度管理に注意する。適当な塩酸濃度の巾も狭いから、古い塩酸を混ぜない方が成績にむらがない。その他は冷蔵期間40日以上の場合に準ずる。
冷蔵期間25日以上30日未満の場合には、液温を47℃、浸漬時間8-9分とし、その他の条件は上に準じ、孵化の斉否および卵の潰れ具合によって浸漬時間を加減するのが安全であろう。
大西(762)は、冷蔵期間30日で支115号×日122号を塩酸濃度20%(15℃比重1.10)、液温45℃で浸酸する場合には、産卵後24℃、35-50時間目または22℃、45-60時間目に5℃に冷蔵し、浸漬時間11分が最もよく、これ以下の浸漬時間では孵化歩合が低下し、また産卵後の保護温度が26℃の場合には、上記に比べて常に成績が悪かったと云っている。保護温度が高いと冷蔵適期の巾が狭いためであろう。冷蔵期間の極端に短かい場合には、出庫後浸酸までの時間を長くする方がよいと云う成績もあるが、冷蔵30日では寧ろ早く浸酸する方がよかった(761)。
短期冷蔵浸酸においては、蚕品種による浸漬成績の相違が長期冷蔵浸酸の場合よりも明瞭である(第140、141表)。
短期冷蔵浸酸がむずかしいと云われている豊年×研白についての、蚕糸試験場新庄原蚕種製造所と山形県蚕業試験場との協力試験成績があるが、それによると、産卵後25℃、36時間目および48時間目(18時基点)に5℃に冷蔵し、30日で出庫、2時間後に15℃比重1.10、液温48℃の浸酸を行なった処、新庄と山形とで反対の傾向を示すような結果が得られている(第142表)。
| 母体別 | 冷蔵時期 (時間目) |
浸漬時間 (分) |
新庄 | 山形 | ||
| 初発3日 孵化歩合 (%) |
初発5日 孵化歩合 (%) |
初発3日 孵化歩合 (%) |
初発5日 孵化歩合 (%) |
|||
| 日 母 | 36 | 5 7 9 |
55 79 78 |
93 95 92 |
96 95 83 |
97 95 84 |
| 48 | 5 7 9 |
65 75 74 |
96 93 92 |
94 92 88 |
98 95 89 |
|
| 支 母 | 36 | 5 7 9 |
31 53 55 |
95 98 95 |
81 95 94 |
99 98 96 |
| 48 | 5 7 9 |
28 36 34 |
97 96 95 |
87 83 88 |
96 98 99 |
|
即ち、新庄の初発3日孵化歩合をみると、浸漬時間9分ではやや落ちるようであるが、浸漬5分の孵化歩合が最も低い。冷蔵時期36時間と48時間とを比較すると48時間が劣る。これらの傾向は支母体に特に顕著である。また初発5日孵化歩合は何れも90%以上を示しているのであるから、初発3日孵化歩合の悪いのは蚕種の越年化が進んでいたか、浸酸刺戟が不足であったかのように思われる。
これに対して山形においては、初発3日孵化歩合の傾向が日母体と支母体とで相違し、日母体では浸漬5分の孵化が最もよく、浸酸時間の長いほど孵化歩合が低下している。冷蔵時期36時間と48時間との間にも、初発3日と5日との間にも孵化歩合の差が少ないから、新庄におけるような、越年化の進行や浸酸刺戟の不足を思わせるような傾向はない。支母体の初発3日孵化歩合はやや劣っているが、浸漬時間の長短による差も、初発3日と5日との差も共に新庄に比べて著しく小さい。
新庄と山形とにおけるこれらの違いは冷蔵浸酸の成績を検討する上で重要な問題を含んでいるが、その分析は行なわれていない。
新庄および山形の成績を通じ、36時間目冷蔵のものの浸酸抵抗性が48時間目冷蔵のものに劣らないことも注目される。これからみると、初発3日孵化歩合の悪かったのは、あるいは浸酸感受性の低いためではないかとも考えられるが、この点も究明されていない。
C 冷蔵浸酸種の再冷蔵
冷蔵浸酸種を冷蔵する必要のある場合には。浸酸後25℃で12時間頃か、または48時間以上たったものを中間温度(15℃)に6時間ぐらいおいてから行なう。浸酸後12時間以後48時間まで、特に24-48時間の間に冷蔵すると白ハゼ卵が発生し易い(473,673,729,1150)。他所で浸酸して送られてきた冷蔵浸酸種は、浸酸日を確かめた上で、危険時期を避け、中間温度を径て冷蔵する注意が必要である。
この白ハゼ卵は温度の激変の影響で起こるものであるから、上記危険時期の蚕種でも、15℃に18時間ぐらいおいてから冷蔵すると被害が軽減する。冷蔵温度としては、2.5℃の方が蚕種は長持ちするが(胚の時期によって異なる)、白ハゼ卵の出る危険は多い。冷蔵浸酸種は、普通には、長期冷蔵の必要はないのであるから、5℃に冷蔵する方が安全である。白ハゼ卵の発生を防ぐためには、冷蔵前後の温度差を12.5℃以内にする必要があると云われているから、15℃の中間温度を通したものでも、2.5℃に冷蔵するとこの温度差の限度になる。
白ハゼ卵は短期冷蔵浸酸種まりも長期冷蔵浸酸種の再冷蔵によって発生し易い。沓掛・黒岩(478)は、冷蔵によって白ハゼ卵の出易いのは、漿膜細胞のカタラーゼ作用が盛んになりつつあるか、または盛んな時期に当ると云っており、生理的なある転換期の卵に起り易いものと思われる。
白ハゼ卵(白死卵)と云うのは、漿膜が破れて一部に集合してしまい、卵の内容が外部から白くみえる卵の総称であるから、原因の異なるものが多く含まれている。冷蔵浸酸においては、浸酸そのものによっても白死卵の生ずることがあり(657,658)、これは出庫後、中間温度(20-25℃)を経過せずに、直ちに浸酸した場合に多い。冷蔵浸酸種の再冷蔵による白ハゼ卵については、上記のほかにも多くの研究がある(680,1061,1062,1093,1094,1098,1099,1105,1120,1121)。
冷蔵浸酸種の再冷蔵は、実用上、長期間の必要はないのであるが、10日間の冷蔵によっても、5日間に比べて孵化歩合が悪かったと云う報告がある(690)。しかし、これは浸酸後再冷蔵までの時間が長いほと差が顕著になるのであって、12時間以内の冷蔵では差がない。
功刀(458)は、産卵後25℃、48時間目に5℃に冷蔵し、35日で出庫、浸酸(比重.10、液温47.8℃、5分)した後、なるべく早く乾かし、乾き次第5℃に再冷蔵(中間温度を通さず)しておけば、その後35日間に亘り、必要に応じて出庫、催青することができ、この間に孵化歩合が低下することはないと報告した。
これは地方蚕業試験場協力試験(73)、その他(797)で取上げられ、催青時に出庫、浸酸した冷蔵浸酸種に比べて孵化歩合に大差なく、稚蚕減蚕の僅かに多いことはあったが、壮蚕の減蚕歩合および繭質には差のないことが認められた。角田・西垣(882)も類似の試験を行なっている。
D 浸酸奏効の判定
浸酸後1日たつと、浸酸の効いた蚕種は無浸酸のものに比べて水引きが深く、即時浸酸種では卵色も幾分淡くみえる。水引きの強過ぎるのは浸酸刺戟の強過ぎた卵で、孵化がおくれたり、死卵になったりする。
三浦(547)は即時浸酸種について次ぎのように述べている。但し、浸酸後の保護温度75゚F(23.9℃)内外の場合である。
刺戟弱く、浸酸の効かなかった卵
12時間後 卵色 黄、卵面膨隆す
24 〃 〃 褐、 〃
刺戟適度で、浸酸の効いた卵
12時間後 卵色 黄、卵面水平に近い
24 〃 〃 淡褐、卵面水平または中央僅かに低い
刺戟過度の卵
12時間後 卵色 黄、卵面僅かに水引きを生ず(殆どが死卵になる)
24 〃 卵面に水引きを生ず(殆どが死卵になる)
卵黄細胞の脂肪性顆粒の分散状態によってもわかるが(839,926)、実用的で確実なのは、浸酸後25℃に保護し、即浸種はまる2日後、冷浸種はまる3日後に解剖して、胚に付属肢が発生しているか否かを調べる方法である。付属肢の有無をみるだけであるから、生体解剖で十分である(第155、156表)。
盛次(593,594)は、卵汁のpHの変化によって浸酸の奏効が早期に判定できると発表した。無浸酸の蚕種のpHは6.4-6.6であるが、浸酸を施すとこれが一旦6.2ぐらいまで下かった後、再びもとの値に回復する。早く低下して早く回復する蚕種ほど孵化歩合がよいと云う。例えば、日7号×支106号を用い、産卵後80゚F(26.7℃)、20時間目に、塩酸液の比重を違えて、液温115゚F(46.1℃)、5分間の浸酸を施した場合のpHと孵化歩合とは第143表の通りである。
| 塩酸比重 | 浸酸後のpH | 孵化歩合(%) | ||||
| 3時間後 | 12時間後 | 22時間後 | 42時間後 | 総孵化歩合 | 最多2日孵化歩合 | |
| 無浸酸 1.060 1.070 1.075 1.085 1.100 1.115 1.130 |
6.4 6.4 6.2 6.2 6.2 6.0 5.4 4.6 |
6.4 6.2 6.2 6.2 6.2 6.0 5.2 4.2 |
6.4 6.2 6.4 6.4 6.3 6.2 5.6 4.2 |
6.4 6.4 6.4 6.4 6.4 6.2 5.2 4.0 |
0 91 97 99 96 60 0 0 |
0 74 92 97 89 54 0 0 |
方法は卵0.5gに対し0.5ccの蒸溜水を添加し、乳鉢で磨砕し、直ちに試験紙を浸してpHを測定する。脱酸を完全にしておかなければならない。冷蔵浸酸の場合にも適用できると云う。永井(625)、浜(148)も浸酸後のpHの変化を報告している。
河合(348)は即時浸酸後の酸化還元電位の変化をみているが、浸酸奏効の判定には結び付けていない。
戸谷・早坂(1070)はニンヒドリン、ミロン試薬、ネスラー試薬、デラフィールド・ヘマトキシリンなどによる卵殻の呈色が浸酸卵と無浸酸卵とで相違することを認めたが、現在の処では、浸酸の強弱までは判別できず、浸酸奏効の判定にも用いられない。
4 浸酸および浸酸後の冷蔵による蚕種の障害
蚕種の浸酸および冷蔵によって畸形蚕の生ずることは古くから知られており、これには遺伝的な性質に基ずくものと、高温、低温または塩酸刺戟の影響による単なる発生の異常とがある。福田・河野は後者について多くの研究を発表している(119,121,122,124,126,128,448-450)。それによると、胚発生上、異常温度によって畸形の生じ易い四つの時期がある。不越年性卵を0℃に5日または20日間冷蔵し、幼虫および卵内幼虫(青み卵になって孵化し得ないでいる胚)の畸形率を調べた結果について云えば、これらの4期は次ぎの通りである(産卵後の保護温度24℃)。
第1期 7時令を中心とする時期
第2期 20時令 〃
第3期 50時令 〃
第4期 100時令 〃
第1期は分裂核の分裂期(Stage2に当る)、第2期は小判形期(Stage4)からダルマ形期(Stage5)に至る間、第3期は胸肢突起が明かに認められ、腹肢突起の生じ初める頃(Stage17)、第4期は反転直前から反転後半に至る背面形成中の時期(Stage21)であるが、生じる畸形の種類が各々の時期によって異なっている。
第1期の冷蔵によって生じるのは、体節の不整、捻転、背面または腹面の融合、胸肢または腹肢の位置あるいは数の異常、尾角の数や位置の異常、瘤状突起の形成、体の1部の欠損など、不規則な畸形である。第2期に生ずるものは体節の前後融合による体節数の減少が特徴で、これに第1期の処理でみられるような畸形が混じている。第3期のものは左右の胸肢や腹肢が服面中央で各節ごとに接近あるいは合体する。第4期の畸形は背面の融合が特徴である。
41℃の高温に5時間接触させても上と同じ4期に畸形率が高い。
越年性卵を冷蔵した後、高温に戻して休眠させ、翌春孵化したものを調べ、あるいは冷蔵後に浸酸を施して孵化させてみると、第2期が30時令、第3期が60時令にずれている。また20時令で加温浸酸あるいは常温浸酸を施した卵においては、加温浸酸区は30-36時令、常温浸酸区は30時令が第2期に当り、胚を調べてみても、加温区では30時令が小判形、36時令がダルマ形てあり、常温区では30時令がダルマ形であった。
即ち、即時浸酸種の冷蔵適期は、畸形発生の面だけからみても、単なる卵令や浸酸後の時間だけによっては決定できないことがわかる。適期は胚の発生段階によってきまるのであるから、加温浸酸と常温浸酸との違いばかりではなく、浸酸の強さ、時期、浸酸後の取扱いによって冷蔵結果の相違することは当然であろう。
大村(754)の報告した畸形蚕は無処理の越年種を掃立てても発現する遺伝性畸形であるが、即時浸酸によって特に発現率が増大すると云うから、福田らの第2期頃の感受性が高いものと思われる。
梅谷(1108,1110)も浸酸によって畸形の発現する系統について報告しているが、産卵後20時間目(ダルマ形直前)の浸酸によって最も多く発現し、35時間(ダルマ形期)までが発現期である。浸酸後直ちに25℃で催青したものに比べ、15℃に1日または30℃-35℃に1日おいてから25℃に移したものには発現が少ないと云う。
その他、浸酸による畸形の発現については多くの報告があり(26,251,252,392,393)、胚盤形成期(Ⅵ2Ba)を一つの危険期としている。
中国における混精交雑の試験の中で(Ⅵ1脚註)、蚕種の孵化が悪く畸形の多かった場合が余剰精子の影響であると論ぜられている。この場合の材料蚕が即時浸酸種であること、および畸形の形態から考えると、われわれの普通にみている浸酸畸形と同じもののように思われるが、その論議においては浸酸畸形には言及されていない。
框製にすると越年種も浸酸種も孵化が良好であるのに、バラ種の浸酸種に限って異常死卵が発現し、孵化歩合の低下する系統がある(480)。非常に特異なことは、バラ種台紙に付着したままホルムアルデヒド2%液に2分間浸し、約30分後に洗落し、少量ずつ寒冷紗に包んで浸酸すれば死卵の発現を防止できるが、ホルマリンで処理したものも、大量を一度に浸酸すると矢張り死卵がでる。卵細胞質に異常のあるために起こり、母親遺伝をする性質であると云うが(481)、浸酸の方法によって死卵の出方の相違する機構は明かにされていない。
仲野(657)は、浸酸成績の向上のためには、孵化歩合ばかりでなく、不孵化卵の検討の必要なことを述べている。これは非常に大切なことであるが、不孵化卵をただタト見的に早期死卵、催青死卵、白ハゼ卵、浸酸無効卵などに区分して、その多少を調べるだけでは不十分である。催青死卵は浸酸が弱過きても、強過ぎても出るのであるから、その発現の機構に触れた検討が必要である。白ハゼ卵、早期死卵、浸酸無効卵などについても同様で、その死因、原因の分析に進まなければ意味がない。この点に関して興味のあるのは福田・河野(120,123,137)の研究である。
浸酸の時期が適期を過ぎていた場合、浸漬時間の不足な場合などに生ずる浸酸無効卵を解剖してみると、胚のある程度発育しているものが多いが、その中に、頭部は殆ど休眠胚のままでありながら、尾端の発育の進んでいるもの、頭部は臨界期またはそれ以前の形にとどまっていながら体の後半部または尾端の数節が器官形成期に入っているもの、または反転を終えても頭部の発育の著しくおくれているものなどが縷々発見される。越年卵性蛹または不越年卵性蛹を除脳したり、不越年卵性蛹に食道下神経節を移植した場合に、化蛹してから産む越年性卵様の卵において、休眠要因(休眠ホルモン)の不足のために異常な発育をする胚が認められるが、上記の異常胚はこれとよく似ているから、休眠要因またはこれに関係のある物質が不十分な量で存在するために起こる発育異常と考えられる。浸酸刺戟の不十分な卵をある期間冷蔵するとこれらの異常胚の出方が減少するが、これは冷蔵によって活性化が補われるためと考えられる。異常胚において体の後半部の発育が進むのは、休眠要因または休眠解除後の代謝系に対する胚の反応に頭尾勾配があるためであろうと云う。
冷蔵浸酸の場合、出庫後、中間温度を経ずに直ちに高温の塩酸に浸酸すると蚕種に被害があるが、河野(451)は40日間冷蔵した蚕種に、25℃で出庫直後、10、30および60分後に15℃比重1.110、
液漫48℃の浸酸を施こし、その被害を調べた。これによると、浸酸時間がある程度以上長くなるに連れて、どの時期の浸漬においても孵化歩合が低下したが、出庫後浸漬までの時間の短かいほどこの低下が顕著であって、孵化歩合95%以上を示した浸漬時間は、出庫後10分の浸漬では4分、30分後の浸漬で5分、60分後の浸漬では7分がそれぞれ限度で、浸漬時間がこれより長くなると孵化歩合は著しく低下した。処が、出庫直後の浸漬は却って被害が少なく、95%以上の孵化の得られた浸漬時間の限度は6分間てあった。出庫直後の浸漬がよかったと云うことは、普通には考えられないことで、何か特別の条件があったものと思われるから、その原因の究明が望まれる。なお、この場合の被害卵は、浸酸後間もなく漿膜と卵殻との間に間隙が生じ(XI1
E)、大部分のものが反転後から青み卵までの間に死卵になった。
冷蔵浸鼓種の再冷蔵による白ハゼ卵についてはⅦ3Cにおいて述べた。
5 随時孵化および冬掃き蚕種の浸酸
A 随時浸酸
随時孵化は、蚕種の増殖のための急な必要に応じて、休眠中の蚕種を孵化させたいときに非常手段として行なわれるが、成績が不定で、蚕品種、産卵後の保護環境および日数によって結果が著しく相違する。浸酸を主とし、これに低温処理その他の方法が併用される。
水田・佐藤(585)は、1)浸酸の反覆、2)冷蔵浸酸、3)浸酸冷蔵、4)冷蔵浸酸冷蔵、5)浸酸冷蔵浸酸、6)冷蔵浸酸の反覆、7)浸酸冷蔵の反覆などの方法によって、現行蚕品種における随時孵化成績を比較したが、従来から云われているように(547)、浸酸冷蔵浸酸法が最もよかった。
日122号、日124号、豊年、支122号、支122号(太)、支124号および研白についての試験結果を綜合すると、この処理に必要な最小限の冷蔵日数は第144表の通りであった。
| 希望条件 | 産卵後処理までの25℃保護日数 | ||
| 10日間 | 30日間 | 50日間 | |
| 初発3日孵化歩合50%以上を得るための冷蔵日数 初発5日孵化歩合50%以上を得るための冷蔵日数 初発5日孵化歩合90%以上を得るための冷蔵日数 |
20-30 20 20-30 |
20-40 20 20-* |
20-* 20-* 30-* |
浸酸冷蔵浸酸法は浸酸後ある期間冷蔵し、再浸酸を施す方法であるが、期日が切迫して冷蔵処理を行なう余裕のない場合には、浸酸反復法がよいと云う。これは15℃比重1.10、液温48℃で原種は15分、交雑種は20分間の浸酸を行ない、25℃に1日保護した後、同じ浸酸を繰返えした。産卵後25℃、35日の支122号、支122号(太)および支122号×日122号にこの方法を施した場合の初発5日孵化歩合は34-51%であった。
星沢(227)は卵令30日前後の交雑種を60゚F(15.6℃)に1日おいた後、40゚F(4.4℃)に20-30日間冷蔵し、比重1.10、117゚F(47.2℃)、17分の浸酸を行なって良好な孵化歩合を得ている。
東畑・功力(1055)は、産卵後25℃に保護中の蚕種に液温47.8℃、比重1.10、10分間の浸酸を施した後、これに酸素接触を行ない、酸素接触の有効なことを報告している(第145表)。酸素接触は酸素90%の瓶内に蚕卵を密封するのである(141)。
| 蚕品種 | 産卵後処理までの 25℃日数(日) |
浸酸のみによる 孵化歩合(%) |
酸素接触併用による孵化歩合(%) | ||
| 接触3日間 | 接触4日間 | 接触5日間 | |||
| 日9号 | 20 40 70 80 120 |
35 12 46 14 23 |
69 - - 21 43 |
52 40 58 - 52 |
- - - - 50 |
| 日111号 | 40 20 40 |
18 67 48 |
54 67 - |
39 66 - |
- - 60 |
| 支17号 | 40 20 40 70 80 120 20 40 |
71 84 73 65 74 29 59 82 |
84 84 - - 81 54 80 93 |
81 86 81 84 - 55 - - |
- - - - - 55 - - |
| 支106号 | 20 40 |
31 21 |
45 - |
66 - |
- 61 |
| 欧19号 | 70 80 120 |
21 16 10 |
- 26 18 |
41 - 18 |
- - 22 |
石川ら(256,257)は、15℃比重1.100、液温48℃の高温浸酸、40℃付近の高温接触、-10℃以下の低温接触および比重1.126、液温28℃の低温浸酸を組合わせる方法によって、随時人工孵化の成績を向上させることができると云っているが、他の研究者によって確認されるに至っていない。コオロギにおいては、越年性卵を-16.5℃に20分間接触させると、5℃に十数日間接触させたのと同じ程度に活性化したと云う実験(オーストラリアにおける)がある。このコオロギの卵は非常に活性化し易く、また0℃以下の温度において目立って活性化の進む性質があるので、蚕卵とは著しく趣きを異にしてはいるが、Ⅷ3Aにおいて述べたこととも考え合わせると、暫く実用の問題をはなれて、違う角度からこの方法を検討する必要があるように思われる
B 冬掃き蚕種の浸酸
未だ活性化の十分でない越年種を、増殖用として冬または早春に掃立てるためには、浸酸を施して活性化を補う必要があるが、その成績は蚕品種および蚕種の経歴によって相違する(第146表)。
| 研究者 | 採種時期 (場所) |
浸酸時期 | 蚕品種 | 浸酸 | 孵化 歩合 (%) |
備考 | ||
| 塩酸比重 | 液温 | 浸漬時間 | ||||||
| 星沢(227) | 6月 (東京) 記載なし(福島) 〃 〃 〃 〃 〃 (松本) 〃 (福島) 〃 〃 〃 〃 〃 (松本) |
2月5日 11月30日 〃 〃 〃 2月3日 〃 〃 2月7日 |
支5号 欧17号 支14号 支15号 支105号 欧17号 支14号 支15号 支105号 |
1.10 〃 〃 〃 〃 1.08 〃 〃 〃 |
115゚F 118゚F 〃 〃 〃 115゚F 〃 〃 114゚F |
5 7 7 7 5 7 7 7 5.5 |
86 60 60 80 95 82 85 90 95 |
31/X-30/XI 40゚Fに冷蔵 〃 〃 4/XI-30/XI 40゚F 8/I-3/II 〃 〃 〃 〃 〃 〃 〃 8/I-7/II 〃 |
| 前田(502) | 6月 (三重) 〃 〃 8月 〃 〃 〃 |
1月17日 〃 〃 〃 |
支14号 欧16号 日110号 支106号 |
1.10 〃 〃 〃 |
46℃ 〃 〃 〃 |
16 16 16 8 |
75 63 94 77 |
孵化歩合は最多2日 |
| 塚越(1085) | 7月 (山形) 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 9月 〃 〃 〃 〃 〃 |
12月8日 〃 〃 〃 〃 〃 〃 12月23日 〃 〃 〃 〃 |
日7号 日110号 分離白1号 支106号 支15号 支16号 欧18号 日7号 支106号 日7号 日110号 支106号 |
1.10 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 〃 |
117゚F 〃 〃 〃 〃 〃 〃 115゚F 〃 〃 〃 〃 |
5 7 12 6 10 6 10 8 5 8 8 4 |
51 41 46 43 46 35 42 44 48 60 58 58 |
孵化歩合は初発5日。 蚕種の保護温度(平均) は12月上旬4.4℃、 同中旬4.6℃ |
佐藤ら(824)は、新庄の自然温度で保護中の最近の蚕品種について、比重1.10、液温47℃の浸酸成績を比較した。浸漬時間は3、5および7分であったが、最も孵化歩合の高かった場合は第147表の通りであった。
| 蚕品種 | 1月21日 | 2月1日 | 2月15日 | 3月1日 | 3月15日 |
| 日122号 | 0-29% 3-56 |
0-52% 5-76 |
0-66% 3-81 |
0-73% 3-79 |
0-64% 3-66 |
| 日124号 | 0-13 7-44 |
0-31 7-41 |
0-55 0-59 |
0-50 3-62 |
0-45 5-75 |
| 豊 白 | 0-45 3-58 |
0-53 5-77 |
0-61 3-83 |
0-77 7-87 |
0-86 5-90 |
| 研 光 | 0-49 3-60 |
0-38 7-76 |
0-63 7-82 |
0-64 5-81 |
0-60 5-86 |
| 支122号(太) | 0-57 3-78 |
0-81 5-86 |
0-92 5-93 |
0-85 3-87 |
0-95 3-92 |
| 支124号 | 0-77 3-86 |
0-80 5-87 |
0-89 3-92 |
0-91 5-93 |
0-94 3-93 |
| 銀 白 | 0-42 5-76 |
0-60 7-81 |
0-60 7-90 |
0-72 7-87 |
0-70 5-92 |
| 春 白 | 0-44 3-58 |
0-61 7-80 |
0-53 7-86 |
0-71 5-86 |
0-73 5-88 |
2月下旬-3月上旬の催青用には、比重1.10で支那種には100゚F(37.8℃)8-20分、または105゚F(40.6℃)、4-8分、日本種には100゚F、10-30分、または105゚F、15-25分の浸酸がよかったと云う成績もある(215)。
6 人工越冬
人工越冬法の実用的な価値は少ないが、冷蔵浸酸法の冷蔵時期および冷蔵期間、越年種の活性化の遅速などの問題を考える上に重要な関係がある。
越年化の進んだ卵(第51、115表)および産卵後1日以内の卵(第95表)の人工越冬については先に述べたが、その中間の時期の卵については藤原(102,103)の報告がある(第148表)。
| 項目 | 低温 処理 日数 (日) |
蚕品種 | |||||||||||
| 支115号 | 支122号 | ||||||||||||
| 処理開始時期* | |||||||||||||
| 24 | 36 | 48 | 60 | 72 | 84 | 24 | 36 | 48 | 60 | 72 | 84 | ||
| 孵化歩合 (%) |
30 45 60 75 |
0 84 92 95 |
17 92 94 94 |
0 50 94 96 |
0 21 93 95 |
0 16 9 92 |
0 6 89 93 |
71 84 82 88 |
91 94 91 95 |
79 93 93 96 |
74 92 92 95 |
54 92 92 93 |
51 93 96 97 |
| 最多発蟻 までの 催青日数 (日) |
30 45 60 75 |
- 27 16 15 |
41 27 18 17 |
- 32 20 20 |
- 32 22 20 |
- 34 22 20 |
- 35 22 20 |
27 16 15 15 |
24 15 15 15 |
27 18 16 15 |
31 20 16 15 |
31 21 18 15 |
35 27 18 17 |
この結果において片目されるのは、36時間目に低温処理を始めたものの孵化が最も早いことである。藤原は、48時間以後に処理を始めたものの孵化がおくれるのは越年化が進んているためであり、24時間目のものも亦おくれるのは、活性化し得る程度にまで発育が進んでいないことと共に、低温に対する抵抗力の弱いためであろうと考えている。最多3日孵化歩合も36時間目から処理したものが最も高かった。
野口(689)は、産卵後20時間目、48時間目および10日目からO℃、2.5℃および5℃で処理した結果を比較したが、2.5℃においては、20時間目よりも48時間目に処理を始めたものの催青日数が短かいのに対し、5℃においては20時間目のものの催青日数が短かかった(第149表)。
| 処理 温度 (℃) |
低温 処理 日数 (日) |
処理開始時期 | |||||
| 産卵後20時間目 | 産卵後48時間目 | 産卵後10日目 | |||||
| 調査項目 | |||||||
| 催青日数(日) | 孵化歩合(%) | 催青日数(日) | 孵化歩合(%) | 催青日数(日) | 孵化歩合(%) | ||
| 0 | 20 40 60 |
- - - |
- - - |
61 50 31 |
0 0 1 |
- - |
- - 0 |
| 2.5 | 20 40 60 |
- 43 19 |
- 19 62 |
50 33 17 |
0 52 83 |
- 43 33 |
- 1 8 |
| 5 | 20 40 60 |
46 28 15 |
0 51 76 |
48 30 18 |
2 74 84 |
64 49 30 |
0 3 14 |
この結果は、冷蔵浸酸の基本問題について述べたのと同様に、卵の抵抗力と越年化の進行との釣合いがこの場合にも関係していることを示すもので、若い卵が活性化し難いか否かを孵化の遅速や孵化歩合の高低だけによって簡単に推定することはできない。
交雑種を産卵後36時間目から5℃で処理した場合の活性化の速さは、両親の平均よりは母体に近いが、父親の影響も明かに認められる(104)。
なお258頁参照。
Ⅸ 催青
催青と云うのは管理した環境の下で蚕種を孵化させることである。蚕種が青み卵(Ⅵ2Bf)
になることも催青と云うが、ここで云う催青は蚕種を青み卵にし、更に孵化させるための保護・管理である。
これまでに述べてきた蚕種の保護・取扱いおよび人工孵化は、蚕種を最良の状態で催青に移し、最高の孵化能力を発揮させるための準備であって、これらの準備なしに、催青だけによって蚕種の孵化を良くすることはできないが、反面また、1年を費した蚕種保護の苦心も催青の失敗によって無になる訳であるから、蚕種の保護・取扱い、人工孵化および催青は一体になって初めて効果を拳げ得るものである。
ただ、催青とこれらの準備過程との間には蚕種の保護、管理について原則的な違いがある。準備過程においては、越年性の管理(Ⅴ1)と云うことが、直接または間接に、常に問題であったが、催青においては、越年性を全くはなれて、発育の管理が総ての問題である。
不越年種または越年性の解消した蚕種は自然状態に放置しておいても孵化するが、これでは一般に孵化が不斉であり、何時孵化するかの予定も樹たない。例えば、越年種を自然温度で保護しておき、点青したときにこれを2分し、一半は25℃に移し、他半は自然温度のままにおいて孵化させると、第150表のような結果が得られた。
| 調査 場所 |
日115号 | 支108号 | 温度差 平均 (自然温度 マイナス 25℃) |
|||||||||||
| 25℃区 | 自然区 | 25℃区 | 自然区 | |||||||||||
| 孵化 日数 (日) |
総孵化 歩合 (%) |
最多2日 孵化歩合 (%) |
孵化 日数 (日) |
総孵化 歩合 (%) |
最多2日 孵化歩合 (%) |
孵化 日数 (日) |
総孵化 歩合 (%) |
最多2日 孵化歩合 (%) |
孵化 日数 (日) |
総孵化 歩合 (%) |
最多2日 孵化歩合 (%) |
|||
| A | 新庄 福島 飯坂 小淵沢 |
4 3 3 3 |
97 97 82 91 |
94 95 81 89 |
4 5 7 5 |
95 88 85 90 |
90 58 61 84 |
5 3 3 4 |
98 94 84 93 |
94 92 84 87 |
4 4 7 4 |
96 97 89 87 |
90 86 67 75 |
-5.5 -6.9 -6.8 -7.6 |
| B | 綾部 明石 |
4 5 |
86 77 |
85 71 |
5 7 |
87 84 |
82 64 |
4 4 |
98 68 |
93 65 |
5 7 |
95 73 |
86 47 |
-6.7 -7.8 |
| C | 四国 熊本 宮崎 山川 |
6 5 10 15 |
72 86 89 73 |
45 73 45 32 |
11 13 16 28 |
69 85 92 83 |
- 38 37 19 |
5 5 8 12 |
80 80 86 80 |
62 71 63 50 |
11 13 14 19 |
81 81 95 82 |
- 36 37 33 |
-7.5 -9.4 -8.2 -6.9 |
これは農林省蚕糸試験場における支場の共通試験成績であるが、蚕種の保護および催青に開する種々な問題点を示している。
試験場所を北から南へ、A、B、Cの3地帯に分けて考えてみると、AおよびBにおいては、冬の間の低温のために、早く活性化した卵の発育が抑えられ、その間におくれたものの活性化が進むので、春暖と共に発育が始まったときの胚が揃っていて、孵化日数が短かく、総孵化歩合と最多2日孵化歩合との開きも比較的小さい。これに対し、Cにおいては、冬の間の気温が高いため、活度化がおそく、また早く活性化したものが発育を始めるので、孵化が不斉になり、孵化日数が長く、総孵化歩合と最多2日孵化歩合との開きが大きい。
新庄の気温が最も高い(25℃との差が小さい)のは意外な感じもするが、これは、暖地においては早くから徐々に温度が上り、気温は、25℃より、遙に低くても、胚の発育し得る最低温度以上には早く達するため、比較的低温の時期に点青卵になるのに対し、寒地においては、発育最低温度に達するのがおそい代りに、その後の気温の上昇が速いことを示すものである。
ここで重要なことは、自然温度の下で孵化の不斉なC地帯の蚕種を、点青以後だけでも25℃に保護すると、総孵化歩合には変りがないが、最多2日孵化歩合が非常によくなり、孵化日数の短縮する現象である。この傾向は、自然温度での孵化の比較的よく揃うA地帯およびB地帯においても認められる。これは、先きに(Ⅵ3Da)、発育程度の揃っていない蚕種を高温で催青すると、発育の開きが大きくなると述べたことと矛盾するようにもみえるが、そうではない。例えば、第103表において、15℃では反転期から催青期までの発育に16日かかっているのに、25℃では4.18日で同じ発育をしていることからわかるように、発育に要する時間のではない。云わば見かけの上の揃いがよくなっただけではあるが、期限を切って孵化を揃える実用技術の上からは見逃がすことのできない現象である。
催青は孵化試験とは違い、ただ孵化歩合が高いと云うだけでは意味がなく、予定の掃立日に予定量の蟻蚕を最も健康な状態で供給する期限付きの仕事であるから、不確定要素の多い自然環境に頼っていては目的が果し難く、温湿度その他環境条件の管理が必要になる。
1 催青の準備
A 予備催青
本催青の計画をきめるまでに結果がわかるように、予備催青を行なって、催青所要日数、孵化の斉否、初発2日あるいは3日孵化歩合などを調査し、本催青の参考にする。予備催青は抽出検査であるから、本催青の蚕種を正しく代表するように、品種、母体、蚕期、保護法別に材料をとって行なう必要がある。
孵化調査についてはⅦ3B参照。
B 催青室および催青用具の準備、消毒
催青室や催青用具に病毒が付着していると伝染の大きな根源になり、その被害は広い範囲におよぶから、消毒は普通の蚕具、蚕室の場合よりも厳重に行なう必要がある。
しかし、催青室は締切って使用するので、木材などに浸込んて技け切らない消毒剤が、補温を始めてから揮発して障害を起こすことがある。消毒剤の種類によっては、使用前によく補温点検しておく必要がある。
新築の催青室が、塗料や断熱材からの有毒なガスの揮発のために使用できなかった例もあるから、新築、改築、修理などの際には材料に注意すると共に、使用の日までに十分余裕のあるように工事を終えて、予備催青をしておくのが安全である。これは、同じく密閉状態で使用する蚕種庫や冷蔵庫についても同様てあって、洗浄が容易で、病菌や薬剤の侵入および結露を防止できる内面構造を考える必要がある。
斎藤(789)は30×30×1cmの朴の木の板の表裏に各種の塗料を塗り、4月の室温で1カ月間乾燥させた後、30×30×40cmの厚紙製の箱の底に入れ、その上に蚕種をのせて上面をセロファン紙で密閉し、前期を24℃、後期を25℃で催青して孵化を調べた。催青着手のときに、水性壁塗料(2回塗り)、ポリニール(2回塗り)およびセラックニス(砥粉目止、3回塗り)では微かに臭気が残っており、油性調合ペイント(2回塗り)およびクリヤーラッカー(砥粉目止、3回塗り)では上記よりも強い臭気が残っていたが、セラックニスおよびクリヤーラッカー区において、1日目の孵化が幾分少なく、3日目孵化の多かった以外は対照区との間に孵化の差がなかった。発蟻して開封したときにも臭気が残っていた。8月に、塗装後の乾燥期間を15日および1週間にして、同様な試験を行なった結果においても被害は認められなかった。従って、これらの塗料は比較的短期間で無害になるらしいと云う。しかし、蚕種庫、冷蔵庫、催青室などでは乾き方も異なるから、十分に注意する必要がある。
バラ種容器や催青容器、障子張りなどの糊に澱粉質のものを用いると、麹黴その他の菌類の繁殖源になる。このため、糊には防黴剤として、糊材料に対して1割(重量で)のベータ・ナフトール(β-Naphtol)を混ぜ、約10倍の水を加えて煮沸する方法(542)、または糊材料に安息香酸を1%加える方法(544)なども提案されたが、その後、黴の繁殖源にならないCMC、ポリビニールアルコール(Ⅳ5Bc)などが糊料として用いられることが多くなった。更に、ボール紙製(1189)、またはプラスチック製のバララ種容器、催青容器、あるいはその兼用器が作られるようになった(459,796,1021)。これらを大量生産によって安価に作り、一度使ったものは再使用しないと云う考え方も進められているが、今後更に改善すべき点も多いように思われる。
催青容器のホルマリン消毒は、普通の燻蒸では、積重ねた容器の内部までガスが滲透しにくいから、低圧消毒器を用いるのがよいと云われている(470)。
補温装置、冷却装置、補湿器、照明器具、温度計、湿度計その他の点検、室内各部の温度分布の調査を行ない、不良個所の修理をする。温度分布は、室内が空のときと、蚕種を収容したあととでは相違するから、その点の注意も必要である。温度は催青に最も大きな影響を及ぼすから、その調節には特にこまかい注意が大切である(Ⅶ1C)。
C 蚕種および蟻蚕の消毒
卵の外面にカビその他の病菌の付着しているおそれのある越年種は、出庫後ホルムアルデヒドの2%液(ホルマリンの約20倍液)に30分間以内浸した後、水洗、乾燥して催青する。催青中の消毒は、初期および末期に行なえば、出庫当時に行なった場合と大差がないが、中期、特に反転期に行なうと孵化歩合が悪く、孵化日数も延びると云う報告があるが(382)、末期の消毒によっても、呼吸障害のために孵化の1日ぐらいおくれることが多い。
古く、小岩井(418)は、ホルマリン20倍液から発散するガス中に催青卵を5時間密閉すると、蚕種が消毒されて蚕作がよくなると唱えた。その根拠は明かでなかったが、山崎(1178)がウイルス病に対する蚕種消毒に関連して小岩井の業績を紹介し、古田(142)も亦最近、30-40℃に温めたホルムアルデヒドガスで20分間処理すると多角体が完全に不活化され、催青卵(孵化前日)に塗布した麹黴病菌は10分間で完全に消毒されること、20分間処理しても蚕種には害のないことなどを報告した。古田の研究は無菌飼育用蚕種の消毒のための小規模な実験ではあるが、古い業績が新らしい角度から再認識されてきたことは注目される。ウイルスの不活化については、なおXI2参照。
催青中の蚕種に対するホルマリンおよびクライトの影響については梁池(1184)の試験が実用上参考になる(第151表)。
| 試 験 方 法 | 供試卵数 (粒) |
孵化に関する調査 | 蟻蚕の生命調査 | |||||
| 容器の処理 | 催青方法 | 孵化卵 (粒) |
催青死 卵(粒) |
孵化歩合 (%) |
供試蚕数 (頭) |
生命時数 (時間) |
指数 | |
| 無処理 | 標準催青 | 20,262 | 20,115 | 147 | 99 | 105 | 108 | 100 |
| ホルマリン3%液 | 浸漬後直ちに 標準催青 |
〃 | 20,018 | 244 | 98 | 115 | 88 | 82 |
| クライト200倍液 | 〃 | 20,001 | 261 | 98 | 126 | 90 | 83 | |
これは、催青容器を溶液に直接浸して消毒を行ない、その中に蚕種を入れて催青を行なった結果であるが、孵化におよぼす影響は殆どなかった。蟻蚕の生命時数はかなり短かくなっている。
大場(724)は麹黴病菌に対する、10%テトライト石灰(テトライトの有効塩素量は45%)による蟻蚕消毒は、26-27℃、湿度90%の条件では、菌が付着してから7時間以内、特に3-5時間に行なうのがよいとしている。
セレサンその他の水銀製剤の使用は今後は減少すると思われるが、前日にセレサン5%石灰を撒布した容器に蚕種を収容して、催青を行なった結果は第152表の通りであった。
| 試験区 | 供試卵数 (粒) |
掃立当日の 孵化卵数(粒) |
孵化歩合 | |||
| 掃立日(%) | 掃立後(%) | 計(%) | 指数 | |||
| 対照区 セレサン5%石灰尺坪2.0g 〃 1.5g 〃 0.5g |
1,855 1,857 1,853 1,851 |
1,811 1,684 1,689 1,759 |
97 90 91 94 |
1 4 4 3 |
98 94 95 97 |
100 96 96 99 |
また、掃立ての際に催青容器にセレサン5%石灰を撒布して蟻蚕消毒を行ない、掃立後、残った未孵化卵の中から青み卵だけを選り出し、もとの催青容器に収容して、特に青み卵に対するセレサン石灰の影響を調べたところ、孵化歩合はセレサンを撒布しないもの95.6%、撒布したもの79.4%であったが、この場合には、掃立日の孵化歩合が、無撒布区77.5%、撒布区80.0%と後者の方が多かったので、最終孵化歩合は前者99.0%、後者95.7%で、大差はなかった。従ってこの青み卵についての孵化歩合の違いには、セレサンばかりではなく両区の孵化の遅速も関係しているように思われる。
硬化病に対する蟻蚕消毒に用いるセレサンは、セレサン5%石灰が最も普通であるが(485,486,624)、セレサンの水溶液を吸収させて作ったPM加工紙で蚕種や蚕座を包んでおいても予防効果があると云う(487)。
同じく水銀製剤であるシンメルで掃立器を消毒する場合には、1,000倍溶液では差支えないが、それ以上の濃度になると被害があり、沈殿物が付着しているものを用いると消毒14日後においても悪影響があると云う(454)。
2 催青条件
A 温度
Ⅶで述べたような取扱いをして丙A-丙Bで冷蔵した越年種は、出庫後3日間15℃で保護した後、20℃以上の温度に移す。この15℃期間を予備催青と呼ぶこともあるが、先きに述べた予備催青とは違い、温度の激変を避けると共に、胚の発育を揃えるために必要な(Ⅵ3Da)催青の前段である。丙A-丙Bで冷蔵したとは云っても不揃いがあるから、この前段処理は必要である。
越年卵性を確実にするためには、25℃前後の温度て催青する必要があるが、催青温度が越年卵性の決定に影響するのは胚に胸肢の発生する頃から以後であるから(Ⅴ1Aa)、出庫後3日間を15℃に保護しても、そのために不越年卵性になる心配は殆どない。冷浸種も浸酸後15℃で前段処理をした後に本催青に移すのがよいと云う説があるが(654)、一般には行なわれていない。催青が適正に行なわれておればこの必要はない。
15℃の後は、目的温度に上げて孵化までを一定の温度で保護する平進法、次第に温度を高めて行く漸進法など、種々な温度管理法があるが、最も普通に行なわれているのは平進法、および催青期間の後半を前半より2-3℃高めにする段階的な方法である(第153表)。
| 蚕種 | 温度および日数 | 湿度 |
| 15℃ → 18~20℃ → 23~24℃ → 25~26℃ | ||
| 越年種 | 出庫後 (2日間) 4日間 以後孵化まで 3日間 |
75~85% |
| 浸酸種 | - - (3日間) 全期間 |
催青温度と孵化歩合および胚の発育速度については既に述べたが(Ⅵ3Da)、最近の品種の25℃平進催青における胚の発育段階を第154一156表に示す。
| 蚕品種 | 25℃催青 | ||||||||||
| 催青開 始の時 |
1日後 | 2日後 | 3日後 | 4日後 | 5日後 | 6日後 | 7日後 | 8日後 | 9日後 | 10日後 | |
| 日122号 日124号 支115号 支122号(太) 支124号 |
15-16 14-15 14-15 15-16 14-15 |
16-17 15-16 16-17 17-18 16-17 |
18-19 17-18 19-20 19-20 18-19 |
20-21 19-20 20-21 20-21 20-21 |
21 20-21 21-22 21-22 21-22 |
22-23 21-22 22-23 22-23 22-23 |
24-25 23-24 24-25 23-24 23-24 |
25-27 24-25 25-27 25-27 25-27 |
27-28 25-27 28-29 28-29 28-29 |
28-29 28-29 初発 〃 〃 |
初発 〃 |
| 日122号×日124号 日124号×日122号 支115号×支124号 支124号×支115号 日124号×支122号(太) 支122号(太)×日124号 |
14-15 14-15 14-15 15-16 14-15 14-15 |
16-17 16-17 17-18 17-18 17-18 17-18 |
18-19 18-19 18-19 19-20 19-20 19 |
20-21 19-20 20-21 21 20-21 20-21 |
21 20-21 21-22 21-22 21-22 21 |
22-23 21-22 22-23 23-24 22-23 22-23 |
23-24 23-24 24-25 24-25 24-25 24-25 |
25-27 25-27 25-27 25-27 25-27 25-27 |
28-29 28-29 28-29 28-29 28-29 28-29 |
〃 〃 〃 〃 〃 〃 |
|
| 蚕品種 | 浸 酸 後 | ||||||||||
| 20時間 | 40時間 | 2日 | 3日 | 4日 | 5日 | 6日 | 7日 | 8日 | 9日 | 10日 | |
| 日122号 日124号 支115号 支122号(太) 支124号 |
6-7 6-7 6 6-7 6-7 |
17-18 17-18 17-18 - 17-18 |
17-18 17-18 18 18-19 18 |
20 20 20 20-21 20 |
21 21 21 21-22 21 |
22-23 22-23 22-23 23 22-23 |
23-24 23-24 23-24 24-25 24 |
24-25 25-27 24-25 25-27 25-27 |
27-28 27-28 27-28 28-29 28-29 |
29 28-29 29 初発 〃 |
初発 〃 〃 |
| 日122号×日124号 日124号×日122号 支115号×支124号 支124号×支115号 日124号×支124号 支124号×日124号 4・2×5・4 5・4×4・2 |
6 6-7 6-7 6-7 6-7 6-7 6-7 6-7 |
17-18 17-18 18 17-18 18 - 18 17-18 |
17-18 17-18 18 17-18 18 17-18 18 17-18 |
19-20 20 20 20 20 19-20 20 20 |
21 21 21 21 21-22 21 21-22 21 |
22-23 22-23 23 22-23 22-24 23 23 22-23 |
23-24 23-24 24-25 24-25 25 24-25 24-25 24-25 |
24-25 25-27 25-27 25-27 25-27 25-27 25-27 25-27 |
27-28 27-28 28-29 28-29 28-29 27-28 28-29 28-29 |
28-29 29 初発 〃 〃 29 初発 〃 |
初発 〃 初発 |
| 蚕品種 | 浸 酸 後 | |||||||||
| 1日 | 2日 | 3日 | 4日 | 5日 | 6日 | 7日 | 8日 | 9日 | 10日 | |
| 日122号 日124号 支115号 支122号(太) 支124号 |
13-14 12-13 12-13 13-14 12-13 |
15-16 16-17 13-14 16-17 14-15 |
17-18 18-19 16-17 18-19 17-18 |
19-20 19-20 18-19 19-20 19-20 |
20-21 20-21 20-21 20-21 20-21 |
21-22 22-23 21-22 21-22 21-22 |
23-24 23-24 23-24 23-24 23-24 |
25-27 25-27 25-27 25-27 25-27 |
27-28 27-28 27-28 28-29 27-28 |
初発 〃 〃 〃 〃 |
| 日122号×日124号 日124号×日122号 支115号×支124号 支124号×支115号 日124号×支124号 支124号×日124号 |
13-14 13-14 13-14 12-13 13-14 12-13 |
16-17 15-16 14-15 15-16 15-16 15-16 |
18-19 17-18 17-18 17-18 17-18 18-19 |
20-21 19-20 18-19 19-20 19-20 19-20 |
21-22 20-21 20-21 21 20-21 20-21 |
22-23 21-22 21-22 21-22 21-22 21-22 |
24-25 24-25 22-23 23-24 24-25 24-25 |
27-28 25-27 24-25 25-27 25-27 25-27 |
28-29 28-29 27-28 28-29 28-29 27-28 |
〃 〃 〃 〃 〃 〃 |
松村らの結果(Ⅵ3Da)からわかるように、催青日数は20℃と24℃とでは約4日半、24℃と28℃とでは約2日違うから、催青室内の温度分布に注意し、蚕種の上下積みかえを行ない、位置による発育の遅速が起こらないように配慮する必要がある。催青期間は卵の大小によっても、幾分相違する。
孵化を促進するために、催青末期の保護温度を27℃ぐらいまで上げることも行なわれているが、温度が高いと孵化した蟻蚕の体力消耗がはやい(第157表)。
| 催青温度 (℃) |
体重指数 | ||
| 掃立直前 | 1日後 | 2日後 | |
| 25 27 |
100 101 |
434 388 |
1,151 957 |
バラ種の催青においては、早く孵化した蟻蚕が2夜包みまたは3夜包みになることが避けられないが、温度が高いと、特にこれらの早く孵化したものの体力消耗が大きいから注意しなけれはならない。
原種の催青温度は25℃以上に上げないことが望ましい。
仲野(659)が、支105号の越年種を、最長期以後、度度24℃、湿度75%で催青し、種々な期間異常温湿度を作用させた結果によれば(第158表)、反転期以前の温度を20℃にするとその間の湿度も低い方がやや良好であるが、反転後の20℃においては高湿度がよい傾向を示した。これに対し、温度を32℃に上げた場合には、何時の時期でも常に高湿度がよかった。仲野は、最長期-反転期24℃、反転期-孵化期25.6℃、湿度全期間75%の催青を合理的催青法と云っている。
| 異常温湿度 | 実用孵化 歩合(%) |
不孵化卵歩合(%) | 経過日数(日・時間) | |||||||
| 温度 (℃) |
湿度 (%) |
接触時期 | 早期 死卵 |
催青 死卵 |
最長期 | 反転期 |
反転期 | 眼点期 |
眼点期 | 全青期 |
全青期 | 孵化期 |
計 | |
| 20 | 25 | 最長期-反転期 〃 -眼点期 〃 -全青期 〃 -孵化期 反転期-眼点期 〃 -全青期 〃 -孵化期 眼点期-全青期 〃 -孵化期 全青期-孵化期 |
93 91 87 81 91 93 87 93 81 87 |
1 2 1 3 2 0 1 2 2 1 |
1 2 1 5 2 2 3 2 3 2 |
5・00 5・00 3・08 3・08 3・08 |
3・00 6・00 6・00 3・00 3・00 |
1・00 1・16 2・00 2・00 1・08 |
2・02 1・10 1・18 2・02 2・10 |
11・02 14・02 13・02 10・10 10・02 |
| 20 | 95 | 最長期-反転期 〃 -眼点期 〃 -全青期 〃 -孵化期 反転期-眼点期 〃 -全青期 〃 -孵化期 眼点期-全青期 〃 -孵化期 全青期-孵化期 |
92 91 85 83 93 89 85 92 93 92 |
1 2 1 2 2 1 2 1 2 0 |
1 3 7 2 1 3 1 2 0 1 |
5・00 5・00 3・08 3・08 3・08 |
3・00 6・00 6・00 3・00 3・00 |
1・08 1・16 2・00 2・00 1・08 |
2・02 1・10 1・18 2・02 2・10 |
10・02 14・02 13・02 10・10 10・02 |
| 32 | 25 | 最長期-反転期 〃 -眼点期 〃 -全青期 〃 -孵化期 反転期-眼点期 〃 -全青期 〃 -孵化期 眼点期-全青期 〃 -孵化期 全青期-孵化期 |
25 7 1 0 92 77 73 93 90 88 |
48 78 79 69 0 4 1 0 1 0 |
25 12 17 30 3 11 24 1 5 5 |
3・16 3・16 3・08 3・08 3・08 |
3・08 3・08 3・08 3・00 3・00 |
1・00 1・08 1・08 1・16 1・00 |
2・02 1・18 2・02 2・02 1・18 |
10・02 10・02 10・02 10・02 9・02 |
| 32 | 95 | 最長期-反転期 〃 -眼点期 〃 -全青期 〃 -孵化期 反転期-眼点期 〃 -全青期 〃 -孵化期 眼点期-全青期 〃 -孵化期 全青期-孵化期 |
48 21 22 20 72 85 79 90 93 94 |
3・08 3・08 3・08 3・08 3・08 |
2・16 3・00 3・00 3・00 3・00 |
1・00 1・08 1・00 1・00 1・00 |
2・02 1・10 1・18 1・18 1・18 |
9・02 9・02 9・02 9・02 9・02 |
||
| 20 22.8 25.6 28.3 |
75 75 75 75 |
5・00 4・00 3・00 3・08 |
5・00 3・00 2・16 3・00 |
2・00 2・00 1・00 0・16 |
2・02 2・02 2・02 2・02 |
14・02 11・02 9・02 9・02 |
||||
| 最長期-反転期24℃、75%、反転期-孵化期25.6℃、75% | 3・08 | 2・16 | 1・00 | 2・02 | 9・02 | |||||
一般に、高温催青は繭質をよくすると云われているが、蚕種に悪影響のあるような高温は収繭量にも繭質にもよくない(624a,580)。従って、化性に影響がなく、蚕種の生理上適当な催青温度は、自然に、23℃-25℃、これより上っても26℃ぐらいまでの範囲内に定まってくる。
催青温度と繭質や蚕作との関係については多くの報告があるが、催青と云う出発点と繭質や蚕作と云う終末とを、中間過程を抜きにして直接結び付けることには、縷々繰返えすように、結果がどちらであっても無理が多い。
絹生産に直接関係のある絹糸腺に問題を絞って考えると、絹糸腺の細胞数は催青中に決定するのであるから(695)、催青温度の高低によってその数が影響されそうに思われるが、実際にはその差が認められない(711)。
有賀(61)は、中部糸腺に縊れの生ずる不結繭蚕の発現と催青温度(25℃および15℃)との関係を調べたが、一定の傾向が認められなかった。但し、高橋ら(909)は不結繭蚕防止の催青温度としては24-25℃がよいとしている。
15℃と25℃と云うような極端な催青温度の比較になると、化性の変化を通しての影響も考えなければならない。例えば、15℃暗催青においては、25℃明催青に比べて繭質は劣るがラウジネス繊維が少ない(679)、蚕児の食下量および消化量が少なく、尿酸の排泄が多い(212)、などは化性変化の影響によることが大きいものと考えられる。
平山(200)は、毎日12時間毎に変温(20℃と30℃および22℃と28℃)させる区、22℃と28℃との12時間毎の変温を催青着手-反転、反転-点青および点青-孵化の間それぞれ続ける区ならびに25℃恒温区を設けて、孵化、収繭量および繭質を比較し、25℃恒温区が総べての点において最もよかったと報告している。
催青中の高温接触については荒木・三浦(55)、鈴木ら(899-901)の試験もあるが、催青何日目とあるだけて、発生段階は明示されていない。なおⅥ3Da参照。
大宮・中島(743)は31℃(孵化歩合60%を目標にする)と25℃(孵化歩合95%を目標にする)との催青を6世代続けて、その影響が後代の繭質その他に現われるか否かを試験しようとした。この種の試験には必らず淘汰の影響が加わるので結果の判定がむずかしいが、概して云えば、後代の形質に大きな影響はなかったと云う。
催青温度が高いと蟻蚕の体色が淡くなる(Ⅵ3Da)。
B 光線
不越年卵性蛾を出さないためには、催青中、1日16時間以上の照明をするのが原則であるが、特に反転後から気管の形成される頃までの照明が大切である。照度は卵面で5lux以上とする。包み紙1枚を通すと照度が約8割に減ることも注意する必要がある。
光の有無は、また胚の発育および孵化を促進したり抑えたりするので、これを利用して孵化をある程度揃えることがてきるが(Ⅵ3Dc)、これを特に大量のバラ種催青に応用したのが熊本県蚕業試験場で始まった改良暗催青法である。
この方法は、点青の前日から催青容器に黒紙を覆って光を遮断し、まる4日間暗黒で催青を続け、5回目の朝、黒紙を除いて一斉に孵化させるもので、発育の進んでいるものも、暗黒のために孵化が10時間ぐらいまでは抑えられるので、発蟻が揃い、2夜包み、3夜包みの害も少ない。しかし、催青卵になると呼吸が盛んであるから、孵化は抑えられていても消耗は大きく、抑制しないものに比べると、蟻蚕の体重(第159表)、絶食生命時数(第160表)、食桑などにその影響がみられる(878,908,1058)。蟻蚕の取扱いおよび掃立てに当ってはこのことを念頭におく必要がある。
| 区分 | 1g頭数(頭) | 100頭重量(cg) | 同指数 | 備考 |
| 当日孵化区 催青卵5℃冷蔵区 改良暗催青区 蟻蚕5℃冷蔵区 2夜包区 |
2,135 2,165 2,205 2,260 2,335 |
46.9 46.2 45.4 44.3 42.9 |
100 99 97 94 91 |
日122号×支115号 76-78゚F、84-89% 2連制 |
| 区分 | 2例平均(時間) | 指数 | 備考 |
| 当日孵化区 催青卵5℃冷蔵区 改良暗催青区 蟻蚕5℃冷蔵区 2夜包区 |
93 90 88 83 69 |
100 97 94 89 73 |
支115号×日122号 78-80゚F、80-84% 2連制 |
暗催青は完全な暗黒で行なわなければ所期の目的を達し得ないから、この点の注意が何よりも大切である。
暗黒にする時期は、実用試験においては、熊本式のように点青前日からの方が、点青期からよりも効果のある例が多い(324,383)。しかし、実験的には、完全な点青卵(Stage27)になってからでよいと云う成績も出ている。
点青の1日前から暗黒にすると、気管形成期にかかるので、化性変化を懸念する意見もあるが、感温期の末期であること、高温催青であることなでから考えて、普通の品種においては心配する必要がないとされている。
点青前日と云う時期の判定は、本催青よりも1日早く催青に着手する調査区を設け、その点青によって行なうのである。
催青中に螢光灯を照射すると孵化が1日ぐらい早くなり、短期冷蔵浸酸種の孵化歩合も向上すると云う報告があり、点灯によって温度が上がるための影響ではないと云われていたが(359,1015,1016,1019)、螢光灯、赤外灯および普通電灯を照射して、その効果を比較した堀田・河端(228)の成績によれば、点青までは無照射区に比べて照射区の胚の発育が速いが、中でも赤外灯の効果が最も大きく、普通電灯と螢光灯とは大差がないが前者が僅かに速く、何れも熱線の影響と考えられる結果であった。点青から孵化までは光線照射区の発育がおくれ、孵化に至ると無照射区は一時抑止されるが孵化が揃い、照射区の孵化は不揃いであるなでのことも、一般の光線照射の場合と同権な傾向で、螢光灯の特異性は認められなかった。紫外線で蚕種を照射した古い成績(188)をみても、その影響は明かでない。
催青室に螢光灯を使うと、紫外線が出て、蚕種に害があるかも知れないと云われたことがあるが、現在では広く催青室に使用されていて、害はない。普通の卵色の卵ならば、殺菌灯を連続照射しても正常に孵化する。殺菌灯の短期照射によって被害のないことは田辺・萩原(1017,1018)も報告している。
催青卵に超短波を照射すると孵化がよくなり、蚕作も向上すると云われ、一時これに関する報告が多く(39,147,312,413,415,416,1009,1076)、人工孵化にも利用できると云われたが(414,1077)、その効果が一般に確認されないまま中絶の形となり、現在ではこの方面の研究は行なわれていないようである。
C その他の条件
催青温度については各所で言及した(Ⅴ1Aa、Ⅵ3Db、Ⅸ2A)。催青温度23℃-26℃の場合には75-85%が適当であるが、孵化の際には十分に補湿するのがよい。
催青湿度は孵化後の蟻蚕の絶食生命時数にも影響し、催青温度25℃の場合に、60%区は75%および90%区に比べて生命時数が短かったが、75%区と90%区との間には差が認められなかった(514)。なお絶食生命時数は絶食中の湿度が高いほど長い(第161表)。
| 催青湿度 (%) |
絶食中 湿度(%) |
日1号 | 支4号 | 欧9号 | 正 白 | 新 白 | 平 均 |
| 60 | 60 75 90 |
100 112 118 |
100 119 124 |
100 108 110 |
100 101 109 |
100 114 113 |
100 112 115 |
| 75 | 60 75 90 |
100 111 107 |
100 117 117 |
100 109 112 |
100 102 103 |
100 107 112 |
100 110 110 |
| 90 | 60 75 90 |
100 108 109 |
100 109 111 |
100 107 114 |
100 106 113 |
100 109 106 |
100 108 111 |
大宮・大河内(744)は、催青温度25℃および30℃の場合における湿度100、75および50%の比較を行なったが、50%区は他の2区に比べて催青死卵が多発し、孵化歩合が劣り、蟻蚕体重軽く、稚蚕期の減蚕も多かった。低湿の悪影響は卵の小さい品種において大きかった。 100%と75%との間には催青死卵および減蚕歩合についての差はなかったが、後者の蟻蚕体重は幾分軽かった。
催青に要する空気量については、炭酸ガス排出量を基にした計算も一応の基準にはなるが(Ⅵ3B)、催青中は、検知管を用いて簡易に室内の炭酸ガス量を測定し、許容量を超えないように管理することが実際問題としては必要と思われる。気圧および気流についてはⅥ3Dd参照。
秋蚕催青中に蚕種を水に浸して冷温を感じさせ、且水分を与えるとよいと云う説があるとのことで、荒木・三浦(56)は、2月5日から33-38゚F 、平均35.4゚F(1.9℃)に冷蔵しておいた前年7月採り越年種を7月18日に出庫し40゚F(4.4℃)内タトの水に1分および10分間浸漬した後に催青したが、別段に効果は認められなかった。
発生の不揃いな蚕種を水につけ、催青卵を浮かせて選別する業者がある。このようにして選り分けた催青卵には、孵化の1日ほどおくれるものが多いが、総孵化歩合には差がない。経過、特に稚蚕の経過が幾分おくれ、稚蚕の減蚕も幾分多いが、他に悪条件が加わらなければ、その後の収繭量および繭質には差のないのが普通である(昭和38年度関東地区蚕業試験場協力試験)。
催青中の蚕種の異常条件に対する抵抗性試験は極めて多いが、催青中のどの時期が強いとか、薬品の何%までは害がなかったと云うような成績はあっても、程度の問題に過ぎないのであるから、催青中は異常な条件には一切触れさせない注意が大切である。またこれらの試験には胚の発育段階を明示していない成績も多いが、数例を示せば、鈴木・山桝ら(898,902,903)の炭酸ガスおよび水、水井・稲熊(573)の呼吸障害、ホルムアルデヒドガス、石炭酸ガス、高温および低温、荒木・三浦(54)のアンモニア、長田(642,643)のメチルクロライド、平尾・五十嵐(199)のフレオン、辻田(1083)のDDT、中西(655)のBHC、大野(765)のアルコールなどがある。
アルコールは実験の場合の卵面消毒に縷々用いられるので、大野の成績の要点を示すと次ぎの通りである。無水アルコールに10分間浸漬した場合の孵化歩合を無浸漬を100とする指数で示せば、即時浸酸種の催青第1日98、第2日(突起形成)90、第3日(反転前)94、第4日(反転後)65、第5日(剛毛発生直前)60、第6日(剛毛発生)79、第7日(点青)93、第8日(催青)93であったが、孵化歩合が0になる温度および浸漬時間から考えると、最も強いのが催青卵、最も弱いのは反転期、ついで点青卵で、冷蔵その他の場合に云われる強弱と一致している。越年性卵について云えば産卵当初が最も弱く、休眠期が最も強かった。
90%アルコールによる成績もこれと大差がない。
野口ら(691)は、催青中にチリー地震による津波に襲われ、催青室に泥が入り卵面の汚れの甚しかった蚕種の孵化が悪かったことを報じているが、毎水や泥をかぶった蚕種の処置については記載していない。孵化したものの飼育成績は対照に比べて遜色がなかったと云う。
D 二化性種を不越年化させるための催青
養蚕の実用上では、人工孵化種を用いるので、二化性種を不越年化させる催青の必要はないが、各種の実験のために不越年性卵の必要なことがある。
以前は、このための催青法を究理催青法と呼んだが、今日の言葉で云えば低温暗催青である。二化性と云っても品種によって不越年化し易さが異なるから (第91表)、同じ処理を施こしても、全部が100%不越年卵性蛾になるとは限らないが、不越年化のための催青条件は次ぎの通りである。
胸肢発生期(Stage17)以後、特に剛毛発生期(Stage24)以後を15℃に保護する。この低温期間は、日数にして10日以上必要である(Ⅴ1Aa)。胚の反転(Stage21)から頭部着色の頃(Stage26)までを暗黒に保つ。極めて弱い光にも感ずるから、完全な暗黒にする。温度は70%以下、特に反転期以後を低湿に保ち、孵化のときだけ明るくし、孵化を促進するために温度を上げ、補湿する。ただし、点青期以後は乾き過ぎると催青死卵が多発するから、50%以下の湿度は避ける。孵化後の飼育条件、上蔟後の保護条件にも注意する。
E 催青条件と眠性の変化
三眠蚕は、一般に、低温暗催青によって発生し易いが(115,406,409,597,803)、不越年化の場合とは異なり、15℃よりも20℃催青に多く、反転までおよび胚完成後の高温明催青は低温暗催青よりも三眠蚕の発生を多くし、その中間の時期においては温度および光線の作用がこれと反対になると云われている。仲野(659)および佐々木(806)も反転期以後の低温催青が三眠蚕を多くするとしている。また不越年化の場合とは反対に、催青湿度の高いほど三眠蚕が多く出る。この三眠蚕の発生に湿度の影響するのは反転前であって、20℃で催青し、後期の湿度を低くした場合に三眠蚕が多く出たと云う報告のあるのは、低湿のために不越年卵性化した蚕の発育経過が短縮して三眠蚕になったのであろうと説明されている(806,807)。
点青卵を5℃で5日間、蟻蚕を10℃で3日間抑制しても三眠蚕が多くなると云う(805)。佐々木はこれら三眠蚕の発生を総べて、不越年卵性化した蚕の発育経過が短縮したために起こるものと考えており、一化性品種には上記の種々な誘発条件を作用させても三眠蚕が発生せず、二化性蚕に限って誘発されるとしている。佐々木は、また三眠蚕の発生は越年種よりも即時浸酸種に多く、冷蔵浸酸種と越年種との間には殆ど差がなく、浸酸後の台紙の乾きの遅速も関係がないと云う(804)。眠性変化には飼育条件の影響も大きい(808)(第88表)。
五眠白は、反転以後点青までを20℃で催青しても(その他の時期は25℃)反転までを20℃にしたものよりも却って四眠蚕歩合が低くかった(714)。この場合、反転から点青までは20℃で4日18時間であったのに対して、催青着手から反転までは20℃で8日10時間かかっているから、五眠白のような一化性蚕においては、発生段階よりも20℃におく期間の長さが影響するのではないかと云う。また、五眠白においては、産卵直後から20日間30℃で保護した越年種には、この期間を20℃および25℃で保護したものよりも四眠蚕の発生が多いようであり催青卵および蟻蚕の冷蔵によっては四眠蚕が増加せず、冷蔵期間の長くなるほど却って減少するような傾向が認められた。
3 催青中の蚕種の冷蔵および蟻蚕の抑制
A 催青中の蚕種の冷蔵
人工孵化種または不越年種を初めから計画的に冷蔵する場合(Ⅷ2C、Ⅷ3C)とは違い、催青に着手してから急に掃立てを延期する必要の生じた場合には、胚に付属肢の生じ初めた頃ならば直ちに5℃に移すと10-15日間は冷蔵することができ(17,50,445,456,790)、これより催青の進んだ時期ならば、そのまま催青を続けて青み卵で冷蔵するのが安全である(580)。催青卵での冷蔵は5℃で1週間以内が普通で、越年種においては、完全な青み卵になり、僅かに走りの蟻蚕の孵化し始めたときに行なうが、不越年性卵ではこれよりやや早く、青み卵初期がよいと云われている。湿度は75-80%を目標にする。
しかし、冷蔵成績は種々な条件によって相違し、また研究者によっても、細目では一致しない結果が出ている。
斎藤(790)は、冷蔵期間15日までの試験を行ない、催青卵よりも反転直後の冷蔵がよく、従来、冷蔵に最も不適当とされている気管成期に冷蔵しても、孵化歩合は催青卵冷蔵に劣るが、飼育成績を考慮すると大差がないと云っている。
これは重要なことで、孵化直前まで発育している催青卵は、生理的に障害を受けていても一応孵化はするのに対し、それより早い時期の冷蔵障害卵は孵化し得ず、冷蔵障害のなかったもの、あるいは障害の軽かったものだけが孵化する結果、孵化歩合と飼育成績とが反対になることも十分に考えられる。しかし、飼育には複雑な要素が加わって、結果を乱すおそれもあるから、飼育成績を考慮に入れて判定する場合には十分注意しなければならない。蟻蚕の強健性(例えば絶食生命時数など)を検討して、今後明かにしなければならない問題である。
北沢(384)は、浸酸種について、気管発生の前後が冷蔵に弱いと云っている。
小針・室賀(395)によれば、完全催青卵になった蚕種を5℃に冷蔵した場合に、孵化歩合付が95%を下らない冷蔵日数は、欧9号×正白が15日、欧9号および支4号×正白が10日、支4号および正白は5日で、品種によって大差があった。
冷蔵に耐える強さは、また卵の大小によっても異なる。小野寺・鈴木(716)が、上蔟の際に尾角を切って出血させたものの産んだ卵(長径×短径1.35mm2)と無処理蚕の卵(1.50mm2)とを5℃で20日間冷蔵した場合の孵化歩合は、前者75.9 (95.6)%、後者87.5
(98.7)%で、後者においては初日の孵化も多かった。但し、材料蚕は五眠白で、括弧内の数字は無冷蔵のものの孵化歩合である。冷蔵中の湿度についての氏らの調査によれば、催青卵を5℃に20日間冷蔵した場合、湿度100%区の孵化歩合は支106号77%、支110号72%であったのに対し、湿度75区においてはそれぞれ77%および75%で、両区間に差がなかった。湿度40区においては孵化歩合が71%および69%で、上記より畿分劣っていた。
目黒(526)は魚を入れた罐の上部に催青卵を納め、5℃で1-3日間冷蔵すると、魚臭によって孵化歩合の低下することを試験した。
催青中の蚕種ではないが、平山・黒沢(201)は、内容2㍑のガラス器中に、互に接触しないようにして蚕種と薬品(溶液は500cc、粉末は30g)とを密封したものを冷蔵し、被害調査を行なった。蚕種は丙Bおよびそれ以前の越冬中のものを主として用い、薬品はホルマリン、テトライト、セレサン、DDTおよびウスプルンであったが、セレサンは殆ど無害、これに次ぐのがホルマリンおよびテトライトで、ホルマリッン(ホルムアルデヒド?)3%液は2.5℃では20日前後、5℃では15日前後で急に被害が顕著になった。テトライト(300倍)は2.5℃ 25日、5℃ 20日で被害が明瞭になった。 DDT粉末の影響は比較的少なかったが、乳剤は温度に関係なく5日前後で明瞭な被害を現わし、20-25日で全滅した。ウスプルンの害は更に甚しかった。
梅津(1125)は、飼育可能な催青卵冷蔵の限度を知るために、5℃、60日間冷蔵の試験を行ない、眉蚕から孵化した15頭のうち5頭、アスコリー88頭のうち4頭がそれぞれ営繭したと報告している。
催青中の蚕種の冷蔵については、なおⅥ3Da、Ⅷ4および第163表参照。
B 蟻蚕の抑制
蟻蚕の抑制温度は50゚F(10℃)がよいと古くから云われているが(57)、小針・室賀(396)によっても確かめられた。これによると、抑制には10℃が最もよく、5℃ならびに7.5℃がこれに次き、蟻蚕の体重の重いものは軽いものより、交雑種は原種より、それぞれ長期の抑制に耐えるが、品種によって著しく異なり、正白×満月、正白×欧9号、欧9号×正白、欧9号、欧9号×支7号、支7号×欧9号および支7号においては、10日間抑制しても、蟻蚕の生存歩合は99%を下らなかったが、満月、満月×正白および正白においては、蟻蚕の生存歩合が99%を下らない抑制日数は5日であった。生存歩合が99%を下らない範囲の抑制口数ならば、掃立て後の発育のおくれは、1、2令中に回復し、壮蚕期の蚕児の体重には無抑制のものとの間に差がなかった。湿度が低いと抑制成績の悪いことを述べてはいるが、適湿を明記していない。75-80%を目標にするのがよい。
吉田(1202)は、10℃で無補湿(湿度79%)、24時間の抑制よりも、25.7℃、 98%の催青室においたものの方が1-3令および4-5令の減蚕が共に少なく、収繭量も多かった例を報告しているが、これは当日孵化の蟻蚕を用いて試験したものであるから、普通に農家の飼育する蚕のように2夜包み、3夜包みの混っている場合には、このような結果にはならないと思われる。
日129号および支129号についての抑制3日間の試験成績によれば(第162表)、抑制したものの径過のおくれは3令までに回復したが、1-3令の減蚕歩合が幾分多かった。
| 蚕品種 | 試験区 | 供試蚕 数(頭) |
経過日数(日・時間) | 1~3令 減蚕歩合 |
飼育中の | ||||
| 1令 | 2令 | 3令 | 計 | 温度(℃) | 湿度(%) | ||||
| 日129号 | 対照 抑制 |
353 353 |
3・23 3・23 |
3・12 3・12 |
4・00 4・00 |
11・11 11・11 |
0 1 |
26.3 26.3 |
79 〃 |
| 支129号 | 対照 抑制 |
441 441 |
3・23 3・23 |
3・12 3・12 |
4・00 4・00 |
11・11 11・11 |
6 8 |
26.3 26.3 |
〃 〃 |
| 日129号×支129号 | 対照 抑制 |
395 395 |
3・11 3・23 |
3・12 3・05 |
3・12 3・07 |
10・11 10・11 |
5* 1 |
26.4 26.4 |
〃 〃 |
| 支129号×日129号 | 対照 抑制 |
434 434 |
3・11 3・23 |
3・12 3・05 |
3・12 3・07 |
10・11 10・11 |
1 2 |
26.4 26.4 |
〃 〃 |
蟻蚕の抑制と催青卵での冷蔵との結果を蟻蚕の体重指数によって比較すると、第163表の通りで、蟻蚕抑制3日は催青卵冷蔵7日に劣る成績であった。
| 抑 制 | 体 重 指 数 | ||
| 掃立直前 | 1日後 | 2日後 | |
| 無抑制 催青卵7日間 蟻蚕3日間 蟻蚕7日間 |
100 98 96 92 |
424 399 392 373 |
1,294 1,052 1,015 934 |
田崎(524)は、掃立てをおくらせる方法として、発蟻前日の蛹、発蟻当日の蛾、即浸卵、孵化前々日の卵、および孵化当日の蟻蚕のうちのどの段階で抑えるのがよいかを試験している。これらの各時期の処理がそれぞれ適正に行なわれたか否か、例えば、何れの段階での冷蔵も総べて5℃で行なったこと、孵化前々日と云えば点青期と考えられるが、この時期の冷蔵が適当かどうか、また4令-化蛹の減蚕が冷蔵日数とは無関係に著しく多いこと、繭および蛾の場合には交尾歩合、産卵数なども考慮に入れなければ比較ができないことなど多くの問題点を含んではいるが、一つの比較として興味がある。(第164表)。
| 冷蔵 日数 (日) |
試験区 | 孵化 歩合 (%) |
催青死 卵歩合 (%) |
不受精 卵歩合 (%) |
減蚕歩合(%) | 繭重 (g) |
繭層重 (cg) |
繭層 歩合 (%) |
|
| 稚 蚕 | 4令-化蛹 | ||||||||
| 10 | 繭 蛾 浸冷 青冷 蟻 |
93 86 95 97 - |
2 5 3 1 - |
4 7 1 0 - |
3 3 3 5 8 |
41 37 48 52 45 |
1.59 1.72 1.63 1.66 1.67 |
33.5 35.3 32.8 32.5 33.1 |
21.1 20.6 20.1 19.6 19.8 |
| 20 | 繭 蛾 浸冷 青冷 蟻 |
70 70 88 88 - |
2 2 0 5 - |
18 23 0 0 - |
10 10 22 28 36 |
49 51 45 58 43 |
1.51 1.52 1.53 1.47 1.54 |
28.0 28.4 30.0 27.8 29.4 |
18.5 18.7 19.7 18.9 19.2 |
梅律(1125)は、蟻蚕を5℃に冷蔵し、飼育し得る限度の冷蔵日数を試験したが、M195×元2の蟻蚕129頭を32日間冷蔵した場合に、生存していた40頭を飼育して5頭が営繭し、国一XLg支109の蟻蚕を同じ期間冷蔵した場合には、80頭を掃立てて20頭が営繭したことを報告している。
鈴木・鈴木(892)は、蟻蚕を5℃に冷蔵中、機械の故障のため、冷蔵庫内の温度が-6℃まで低下した場合の成績を報告しているが、蟻蚕を室温に1昼夜放置したもの、および5℃に5昼夜冷蔵したものとの減蚕歩合(第165表)、繭質並に収繭量(第166表)の比較によれば、特別な被害は認められない。
| 区別 | 保護室の温度 と保護時間 |
掃立蚕数 (頭) |
減蚕歩合(対掃立蚕数)(%) | |||
| 1-2令 | 3令-結繭 | 死 籠 | 計 | |||
| 室温放置区 異常低温区 5℃区 |
14-20℃、1昼夜 6~-6℃、1昼夜* 5℃、5昼夜 |
506 483 496 |
6 6 7 |
5 3 4 |
0 5 1 |
12 15 13 |
| 区 別 | 繭 質 | 対掃立1万頭収繭量 (繭重×健蛹数)(kg) |
||
| 繭重(g) | 繭層重(cg) | 繭層歩合(%) | ||
| 室温放置区 異常低温区 5℃区 |
2.16 2.16 2.27 |
47.7 47.9 49.9 |
22.1 22.2 22.0 |
18.8 18.3 19.7 |
C 蟻蚕の2夜包み、3夜包み
蚕種の保護、人工孵化、催青なとについて述べた問題は総べて孵化歩合を高め、孵化を揃えることを目的にしているのであるが、現実の問題としては、バラ種の催青において2夜包み、3夜包みの蟻蚕のできることは避けられない。
2夜包み、3夜包みの蟻蚕が孵化当日のものに比べて体力の消耗していることはその体重の減少によっても明かであるが(第167表)、保護条件ばかりではなく、卵の大小、健否などによっても消耗に多少がある。これらの蟻蚕は食桑、行動なども不活溌で、初期の発育がおくれるが(第168表)、弱いものは1-3令の間に減蚕になって除かれ、強いものはこの間におくれを回復し、残ったものだけについて比較すれば当日掃きに劣らない蚕作を示すのが普通である。
| 蚕品種 | 保護温湿度 | 孵化当日 | 2夜包 | 3夜包 | 4夜包 | |||||
| 温度(℃) | 湿度(%) | 実数(g) | 指数 | 実数(g) | 指数 | 実数(g) | 指数 | 実数(g) | 指数 | |
| 支115号×日122号 | 15 25.7 |
76.3 77.7 |
0.200 0.200 |
100 100 |
0.191 0.186 |
95 93 |
0.183 0.174 |
91 87 |
0.179 0.163 |
89 81 |
| 銀嶺×秋花 | 15 25.7 |
76.3 77.7 |
0.200 0.200 |
100 100 |
0.191 0.186 |
95 93 |
0.182 0.175 |
91 87 |
||
| 区 別 | 体 重 指 数 | ||
| 掃立直前 | 1日後 | 2日後 | |
| 孵化当日 2夜包み 3夜包み |
100 93 88 |
100 91 76 |
100 88 73 |
4-5令減蚕は、2夜包み、3夜包みよりも却って当日掃きのものに多いことがあるが、これは、2夜包み、3夜包みの区においては弱い蚕が稚蚕期減蚕として除かれてしまうのに対し、当日掃きの区においてはこれが生残り、壮蚕期の減蚕になるのではないかと云う(376)。
2夜包み、3夜包みによって体力の消耗している蟻蚕を更に弱らさないために、蟻蚕配給の手順や方法および掃立てに注意し、落伍するものを少なくし、早く回復させる配慮が大切である。酒井・石阪(791)は、2夜包みの蟻蚕は食桑不活溌であるが、軟葉を与えると当日孵化のものに近い食下量を示すと云っている。
現行品種として、日129号および支129号についての試験成績を第169表に示す。
| 蚕品種 | 試験区 | 掃立蚕 数(頭) |
経過日数(日・時間) | 1-3令減 蚕歩合(%) |
飼育中の | ||||
| 1令 | 2令 | 3令 | 計 | 温度(℃) | 湿度(%) | ||||
| 日129号 | 対 照 2夜包 3夜包 |
353 353 353 |
3・23 3・23 4・01 |
3・12 3・12 3・07 |
4・00 4・00 4・00 |
11・11 11・11 11・11 |
0 30* 2 |
26.3 〃 〃 |
79 〃 〃 |
| 支129号 | 対 照 2夜包 3夜包 |
441 441 441 |
3・23 3・23 4・04 |
3・12 3・12 3・07 |
4・00 4・00 4・00 |
11・11 11・11 11・11 |
6 6 16 |
〃 〃 〃 |
〃 〃 〃 |
| 日129号× 支129号 |
対 照 2夜包 3夜包 |
395 395 395 |
3・11 3・23 4・04 |
3・12 3・05 3・00 |
3・12 3・07 3・07 |
10・11 10・11 10・11 |
5 9 20 |
26.4 〃 26.5 |
〃 〃 〃 |
| 支129号× 日129号 |
対 照 2夜包 3夜包 |
434 434 434 |
3・11 3・23 3・23 |
3・12 3・05 3・05 |
3・12 3・07 3・07 |
10・11 10・11 10・11 |
1 1 6 |
26.4 〃 〃 |
〃 〃 〃 |
これによってみると、2夜包みまでは当日掃きに比べて1-3令減蚕歩合に差がないが、3夜包みになると明かに差がある。3夜包み以上にならないように注意する必要がある。
掃立前の蟻蚕ではないが石井(249)は、掃立後凍霜害その他の事故のあった場合には、春蚕期1令2日目(初晩秋は1日目)までならば、直ちに7℃に冷蔵すれば7日以内は利用価値のある程度の冷蔵ができると云っている。小野・斎田(710)は蟻蚕を2-4日間常温に放置する試験を行なったが、無給桑で放置したものよりも、2回給桑して後放置したものが、減蚕歩合、収繭量共に勝る場合が多かった。
湿度については298頁参照。
1 洗落し
A 洗落しの時期および水温
洗落し作業は蚕種に摩擦、振動、圧迫などの物理的刺戟を与えるので、これによって蚕種の活性化が促進される(394,1103,1118,1119)。
蚕種の活性化に対する洗落しの影響は、前休眠期や活性化の始まりかけた時期のものに大きいが、完全休眠期の蚕種においても再出卵を誘発する。この場合、再出卵として外観的に認められるのは活性化した卵の一部分で、多くのものは再出卵まで進み得ない不完全な活性卵で、発育が臨界期前後で止まり、外見ではわからないが、死卵になって蚕種の孵化歩合を低下させる。
活性化の難易と云う点だけから考えれば、休眠中に洗落す方が概して活性化しにくく、蚕種に対する悪影響が少ない筈であるが、蚕種の休眠中は気温の高い時期であるため、活性化したものが発育して、上記のように再出卵や死卵になるので、却って結果が悪い。
洗落しの適期は気温が水温より低下し初める頃とされているのは、この頃になると、自然の活性化が初まりかけていて、洗落しによってこれが一層促進されるが、気温の低いために不完全活性卵の発育が抑えられるか、自然では抑えられなくても、直ちに5℃に冷蔵して差支えない時期になっていて、人工的に発育管理を行なうことができるためである。もし。洗落し後の保護温度が高いと活性化が進んでいるだけに、胚の発育が早く、蚕種保護に悪い影響をおよぼす。洗落しによる活性化刺戟は個々の場合によって異なるが、5℃冷蔵20日間に匹敵するぐらいの活性化促進作用があるとも云われている。
河合が多くの研究を発表した洗落しによる再出卵は、洗落しの刺戟による一種の人工孵化と考えられるものであるが(Ⅶ1A)、注目されるのは、それらの研究の総てを通して、洗落し水温の低いほど再出卵の発現が多いから、水温15℃または20℃で洗落すのがよいと主張されていることである。
河合(337,338)が、12月2日を第1回として4月15日までの間に12回、毎回5℃、10℃および15℃の水温で洗落した後、自然温度で保護した蚕種(支118号)を4月15日から催青した結果によると、12月2日と15日との洗落しにおいては洗落し水温による孵化の差は殆ど認められなかった。その後、15℃による洗落しの成績がよくなったが、2月15日から4月15日までは再び水温による孵化の差が縮まった。しかし、全般的に15℃区の孵化が勝り、再出卵(洗落し後4月15日までの間に自然に孵化した卵)は5℃区に最も多かった。河合はこの結果を、5℃の水で洗落す場合には、それ迄に蚕種が3.6-6.4℃の平均気温に接触した日数が多いほど総孵化歩合および最多2日孵化歩合が低く、特に3.5℃以下の日数が30日未満で、3.6-6.4℃に接触した日数の最も多い時期に洗落したものの孵化が悪く、その後、3.5℃以下の温度に接する日数が増加するに連れて両孵化歩合ともに良くなると説明したが、これはただ、このときの孵化成績を気温に結び付けて説明したに過きず、結び付けの必然性、およびそれが卵の生理にどのように影響したのかについては検討が加えられていない。
これが活性化の促進だけに原因があるのであれば、洗落し後、直ちに冷蔵して、活性化した胚の発育を抑えれば被害はない筈であるが、支122号を水温2.5℃と20℃とで洗落し、2.5℃と5℃とに冷蔵した別の試験の結果をみても、矢張り20℃の水で洗落したものの孵化がよかった(342,343)。また、2.5℃で洗落した蚕種の活性化が早く、冷蔵中に早く活力が低下するのであれば、5℃よりも2.5℃に冷蔵した方が結果がよい筈であるが、冷蔵温度の影響は殆どなく、寧ろ5℃に冷蔵したものの良い場合さえあった(第170表)。
| 洗落し水 温(℃) |
冷蔵温 度(℃) |
孵化歩合 の区別 |
洗落し月日(%) | ||||||
| 12月27日 | 1月11日 | 1月21日 | 2月1日 | 2月11日 | 2月27日 | 3月11日 | |||
| 2.5 | 2.5 | 総孵化 最多2日 |
25 22 |
31 24 |
49 44 |
42 38 |
34 28 |
62 53 |
58 53 |
| 5 | 総孵化 最多2日 |
28 23 |
36 28 |
43 36 |
57 49 |
40 35 |
73 66 |
68 58 |
|
| 20 | 2.5 | 総孵化 最多2日 |
59 50 |
81 76 |
92 91 |
88 83 |
80 73 |
90 83 |
84 79 |
| 5 | 総孵化 最多2日 |
80 70 |
81 78 |
95 94 |
92 90 |
90 84 |
86 80 |
79 73 |
|
| 洗落さな いもの |
対照 | 総孵化 最多2日 |
92 81 |
89 71 |
95 90 |
91 87 |
87 82 |
92 86 |
84 81 |
| 洗落し の日か ら催青 |
総孵化 | 85 | 94 | 98 | 95 | 92 | 89 | 86 | |
この成績で注目されるのは、普通の洗落しでは考えられないほど孵化歩合の低下すること、および12月27日に既に孵化歩合85.5を%示すほど活性化した蚕種が材料であったことである。河合は、12月27日洗落しのものの孵化が特に悪いことから、洗落しの時期、従って活性化の程度が洗落しの結果に影響するものと考えてはいるが、洗落しと同時に、洗落さないものを同様に冷蔵した成績、および冬期に洗落したものをそのまま直ちに催青した成績がないので、十分な検討を加えることができない。
別の品種について、12月26日から3月11日までの間に6回、水温2.5℃20℃との水で洗落した蚕種を2.5℃に40日間および110日間冷蔵する試験も行なわれたが(345)、やはり20℃での洗落しがよく、冷蔵40日間では1月末から2月末までの洗落しが悪く、最長期を過きていたと云う3月11日の洗落しがこれらの時期の洗落しに勝る場合があった。冷蔵110日間では洗落し時期のおそいほど悪かった。
このように、12月~1月の洗落し成績が悪いので、支122号および日122号を用い、12月10日と1月12日とに水温15℃で洗落し、2.5℃゜と5℃とに冷蔵しておき、5月7日に出庫、催青したものと、12月10日と1月12日とから、洗落さずに2.5℃および5℃に冷蔵し、5月8日に出庫して15℃の水で洗落し後、直ちに催青したものとの孵化を比較した処、催青直前に洗落したものの孵化が明かによかったが、冷蔵温度は、前の試験とは異なり、洗落し時期と無関係に大部分のもので5℃よりも2.5℃がよかった。
これらの結果は、個々の試験に関する限り、統計的な要因分析が行なわれているが、それらの試験結果相互の関係には問題点が残っている。例えば、ある試験においては冷蔵温度の影響が認められるが、他の同じような試験においてはこれの認められないのは何故であろうか。また、第170表において特に孵化の悪い12月27日および1月11日が、第117表において孵化の悪かった時期とぼほ一致しているのをみると、その場合に述べたようなことをも含めて、洗落し水温以外にも何らかの条件がこの成績に関係しているのではないか、と云うことも一応考えてみる必要がある。
河合の試験によって明かにされたことは、低温の水で洗落すと蚕種の活性化が速いこと、低温の水で洗落した蚕種をそのまま自然温度で保護すると、洗落し時期の如何に拘らず孵化の悪いこと、および上記のように活性化の進んだ蚕種を低温の水で洗落すと、自然温度保護ばかりでなく冷蔵してもし孵化の悪いことである。
一方、普通に行なわれている11月下旬から12月上旬にかけての洗落しは、
長い実際上の経験からみて、特別に蚕種の故障の原因にはならないのであるから、この普通の時期に普通の取扱いをする蚕種の洗落しの場合にも、気温と同じぐらいの温度の水は悪いのか、悪くはないが15℃または20℃の水の方がよいのか、は実用上極めて関心の深い問題である。もし20℃の水を使うのがよいのであれば、湯を沸かしてでも水温を上がるべきであろうか、と云うような疑問が蚕種業者の間で話されたこともあるから、この点の究明が望まれる。ただ現在の処では、実際上の問題点と河合の研究の問題点とが少しくずれており普通の洗落しの場合に、湯を沸かしてまで15℃または20℃の水を用いなければならないと云うだけの試験結果は出ていないのである。
催青着手時期になっての洗落しは、実際に行なっている業者もあって、これがよいか否かについては技術上の問題よりも、経営上の問題として検討すべきことが多い。十万・岸本(300)の成績によれば、日122号改×支122号良を12月19日に洗落し、12月20日 4.6℃、1月1日 2.6℃、 3月12-14日に中間手入れを行ない、以後2.5℃に冷蔵して4月16日に出庫したものと、洗落さずにこれと同じ冷蔵を行ない、出庫後に洗落したものとは、孵化歩合が94%、98%、1-3令減蚕歩合3.0%、2.8%、全令減蚕歩合11.6%、9.3%と、出庫後に洗落したものが何れも幾分勝っていたが、繭質および収繭量は冷蔵前の洗落しが幾分よかったと云う。両者に差がなかったと見るべきであろう。洗落しの水温は15℃目標であった。水温を規定しない戸谷ら(1070a)の結果においても差がなかった。
実用上の問題ではないが、普通台紙に産ませてある越年種をバラ種にするためには、2、3月頃に比重1.10、液温100゚F(37.8℃)の塩酸液に15-20分間浸漬した後、水中で洗落し乾燥後40゚F(4.4℃)に冷蔵しておくのがよく、孵化歩合は対照と差がないと云う(欧18号×支16号)(69)。 また、比重1.075-1.100の塩酸液に自然温度で20分内外浸漬してから洗落してもよい。
B 比重選
蚕卵は卵殻の構造の細隙に空気を含んでいるから、正しい比重を測定することはむずかしい。塩水選の場合に比重と云うのは、用いた塩水の比重である。
普通には、比重1.09-1.10(重比重)の塩水で沈む蚕種を除き、次ぎにこれを比重1.05
- 1.06(軽比重)の塩水に移して浮いたものを除き、残りを水洗、脱塩するか、または軽比重の塩水を用いずに、重比重で洗んだものと水に浮いたものとを除去するのであるが、塩水比重の階級を細かくして、階級別に卵の分布を調べると、品種により、作柄により、また蛾区によっても相違があり、1蛾の卵で中心価が二つ以上のことも珍らしくない(1128)。
比重で分けた卵の内容についての勝又(318)の調査によれは(第171表)、水に浮くものには正常卵は極めて少なく、その中の正常卵だけを選び出しても孵化歩合が劣ったが、比重1.09の塩水で沈んだものの半数以上は正常卵で、その孵比歩合も正常であった。しかし、重比重で沈む卵および軽比重または水で浮く卵の内容は試料によって著しく相違しており、河野(446)、牛込・服部(1128)らの成績によれば、比重の特に大きいものおよび小さいものは共に孵化が悪く、減蚕も多かった。来海・門脇(469)は豊光×新玉の越年種を比重で区分して飼育したが、比重1.04-1.09
の間においては飼育成績に大差がなかった。これより比重の大きいものは概して不良であった。健卵歩合は1.08-1.09の卵において高かった。
交雑種の比重はほぼ母体品種に等しい(592)。
C 蚕種の膠着防止
洗落したあとで卵が付着し合うのは累積(重積)卵の場合を除けば、卵の膠着物が水で柔かくなり、これが付着し合って固まるのであるから(Ⅳ5Bc)、洗落しの際に3%ホルムアルデヒドに30-40分間浸漬して、膠着物が水で柔かくならないように固める方法もあるが、確実なのはクライトの200倍(0.5%)液に浸漬して膠着物を除去する方法である。
クライトを使用すると卵殻が溶けて卵に被害があると云われるのは、濃度が高過ぎたか、処理時間が長過ぎたか、または液温の高かった場合で、200倍、10℃、10分間ならば被害のないのが普通である。浸酸の場合と同様に、濃度、液温、浸酸時間に注意して処理すべきものである。クライトは作った溶液を長くおいたり、何回も使用すると効果が落ちる。このような液を用いて効果がなかったからと云うので、新らしい液の濃度座を高めたり、浸酸時間を長くすると被害を受けるから、必要な最少量の液を作って浸漬し、1回ずつで液を更新すると効果が一定する。
クライトには卵面消毒の効果もあるが(XI2)、卵の色が変わると云ってきらわれることもある。
累積卵は、膠着物を固めるホルマリン処理では離すことができない。累積卵の特に多い場合には、篩で分けて15℃比重1.075の塩酸液に自然温度で10-20分間浸漬した後、水中で軽く揉むか、または90゚F(32.2℃)の温湯中に10-20分浸漬し、軽く揉めば離すことができると云うが、水に長時間浸漬して処理することも行なわれている。
洗落しのすんだ卵は水を切り、寒冷紗などの上に薄く拡げて乾かし、付着し合っているものを軽く揉んで離すのであるが、この代りに、水を切った蚕種を、よく乾かした麦に混ぜ、袋に入れて振る方法も行なわれている。麦が水分を吸って早く乾き、同時に麦とのぶつかり合いによって卵が離れ、揉む代りをするのであるが、衝撃が強過きると蚕種に害を与える。
蚕種は洗落すときの掻落し器による衝撃のほか、各種の衝撃によって障害を蒙るから、麦との混合、選り分けなどの操作の間にも、蚕種に対しては、麦粒に対するのとは違った配慮をする必要がある。
第172表に示すのは、小さなボール紙製の筒に越年種を容れて、73 cm の高さから床の上に落とした場合の蚕種の被害であるが、2回落としたものは明かに孵化がおくれ、3回落とすと孵化も悪くなった。
| 孵化 | 落下回数とその時の孵化歩合(%) | |||||
| 0 | 2 | 3 | 4 | 5 | 10 | |
| 初発日 2日目 3日目 総孵化歩合 |
39( 41) 90( 94) 94( 98) 96(100) |
10( 13) 83( 89) 89( 95) 94(100) |
5( 6) 67( 74) 86( 94) 91(100) |
10( 16) 55( 66) 76( 84) 83(100) |
3( 4) 43( 58) 63( 84) 75(100) |
1( 1) 29( 42) 56( 81) 69(100) |
この場合には、自然温度で孵化させて、孵化までに21日もかかったため、被害を受けた蚕種の発育のおくれが大きく拡大されているのであって(第150表)、25℃催青ならば、この差は見逃す程度に縮まっている筈ではあるが、被害を受けることは確かであるから注意しなければならない。この被害は、容器の中で蚕種が撥ねて容器の内壁に衝突すると大きくなるもののようで、容器に蚕種を充満させると、少量の蚕種を入れた場合に比べて落下の影響が少なく、ブリキ罐(写真フィルムの罐)に入れて落とすと、容器自身もはずむので、ボール紙容器の場合よりも被害が大きかった。この点では、寒冷紗張りのバラ種容器は落下による蚕種の被害が少ない。
また蚕種をバラ種のまま落とし、フリキ罐に受けても孵化のおくれや孵化歩合の低下がみられた(第173表)。風選の場合などには注意する必要がある。
| 孵化 | 落下回数とその時の孵化歩合(%) | ||||
| 0 | 1 | 3 | 5 | 10 | |
| 初発日 2日目 3日目 総孵化歩合 |
6( 6) 62( 63) 96( 98) 98(100) |
5( 5) 40( 43) 88( 95) 93(100) |
3( 3) 31( 33) 85( 90) 93(100) |
1( 1) 20( 22) 81( 87) 92(100) |
1( 1) 5( 9) 64( 81) 79(100) |
脱水のための遠心力の限度についてはⅧ2Ac参照。
2 卵量と蟻量
産卵後の蚕種の減耗量は保護環境ばかりではなく品種によっても相違するから(Ⅵ3Db)、一率には云われないが(1128)、笠井(309)は、卵重から蟻量を算出する便宜のために、各種の資料に基ずいて蟻量百分率を求めた(第174、175表)。
| 蚕品種 | 産卵期 (%) |
10月末-11月 (%) |
催青着手期 (%) |
発蟻前日 (%) |
| 日本種 支那種 欧州種 平 均 |
70 67 66 68 |
75 72 70 72 |
79 75 74 76 |
89 86 84 86 |
| 蚕品種 | 産卵当 日(%) |
産卵翌 日(%) |
3日目 (%) |
5日目 (%) |
7日目 (%) |
9日目 (%) |
11日目 (%) |
12日目(%) (発蟻前日) |
| 日本種および日母体の雑種 支邦一化母体の一、二化雑種 支邦一、二化雑種母体の3元雑種 支那種および支母体の雑種 平 均 |
78 76 72 75 75 |
78 76 72 75 75 |
78 76 72 75 76 |
78 77 73 76 76 |
78 77 73 76 76 |
79 77 74 76 77 |
79 78 75 77 77 |
88 87 90 87 88 |
これは、それぞれの時期の卵量から、後日得られる蟻量の割合を示したもので、これを用いて、ある卵量からの蟻量は
蟻量=(蟻量百分率×卵量)/100
必要な蟻量を得るための卵量は
卵量=100×必要蟻量/蟻量百分率
によって計算することができると云うのである。
交雑種の減耗率は母体品種に等しいか、または僅かに少ないと云う(592)。最近の交雑種についての卵重減耗率は第100表参照。
牛込・服部(1128)によれば、発蟻当日の蟻量と出殼重量との割合は第176表の通りである。
| 蚕 品 種 | 蟻量(%) | 出殻(%) |
| 日1号 日1号×支4号 |
88 88 |
12 11 |
| 支4号 支4号×日1号 |
87 87 |
13 12 |
| 支7号 支7号×欧7号 |
86 86 |
13 13 |
| 欧7号 欧7号×支7号 |
85 85 |
14 14 |
| 支8号 (支4号×支101号)×日107号 |
85 87 |
14 12 |
但し、この調査においては、蟻量と出殻との合計を100とし、呼吸、蒸散などによる減耗を考えていないから、厳密には孵化前の卵重に対する割合とは一致せず、また、出殻は卵殻の一部を喰取られた残りであるから、その割合は完全卵殻の重量割合とも一致しない。
洗落し当時の蚕卵1g粒数については第13、14表参照。
3 蚕種の輸送
大量の蚕種輸送には、現在、専用の自動車が用いられているようであるから、輸送中の管理が容易で、事故の発生に注意することもできるが、少量の蚕種を公共の輸送機関によって送る場合には種々な問題がある。
三谷・金井(543)はDDT油剤で消毒した汽車または船舶で蚕種を輸送する場合の影響を考えて試験を行なった。直接DDT油剤に触れた蚕種は100%死卵(大部分催青死卵)になったが、100cm2の濾紙に1ccの油剤を撒布し、内容積990cm3の木箱の底に敷き、その面から3cm離して箱内に収容した蚕種(催青の各時期)は、撒布直後に収容しても、2-10時間の密閉では被害がなく、撒布後1昼夜たってからならば、撒布面に密接させて1昼夜おいても無害であった。蟻蚕を撒布面において密閉した場合には、撒布後14日たってからでも被害があった。
揮発性の薬品、特に有機溶媒(シンナー、ベンヂン、クレオソート、 ク口口ホルムなど)のガスは非常に有害である。
輸送中に受ける影響のうちで最も普通なのは高温によるものである。冬期間は気温が低く、蚕種の輸送に安全な時期のように考えられがちであるが、この時期の輸送では不時高温(Ⅶ1Da)の害の外に、暖房の被害を受けることが縷々ある。寒いときには人の体が冷えているので、ストーブのそばに寄りそってもあまり熱く感じないが、温度を測定してみると、局部的には、真夏でも経験しないような高温に接していることが多い。
例えば、蚕糸試験場で原々種の郵送に用いている1蛾採り100蛾用ボール紙製孔あき箱内の各部に熱電対を取り付け、500Wの電熱器から20cmの位置に、箱の側面を熱源に向けておいてみると、室温14-15℃のときに、約2分間で熱源側の外側は約70℃、箱の内部の熱源側から3cm内方の部分は3分で約30℃、6分で約40℃を示した(862)。
従って、ストーブを真赤にしている室内なとでは、熱源の付近や高温になっている床の上などに不用意に蚕種をおくと、人体にはさほどに感じなくても蚕種は被害を蒙るおそれがあり、郵送中に、郵便局や郵便車の中でこのような状況におかれる場合のあることは十分考えられるが、実際にもこのような被害は縷々あり、実験的にもその状況を再現することができる。
蚕種の郵送箱には空気の流通をよくするために孔をあける習慣があるが、冬期間の蚕種には、呼吸の面からみてその必要がないばかりでなく(Ⅵ3B、Ⅶ1Da)、上記の結果から考えて、十分に包装するのが寧ろ安全であるが、次ぎの実験によってもこれを確かめることができた。
これは、上記100蛾用蚕種箱に普通のように蚕種を収めたものと、この箱を更に、各辺を2cm宛大きくしたボール紙製の外箱に入れ、内箱と外箱との間にはセロファン紙の細片を詰めた二重包装との比較試験である。
2月下旬から4月上旬までの間、5日毎に東京から明石へ蚕種を郵送し、明石ではこれを目然温度で保護して孵化を調べた。蚕種は東京で5℃に冷蔵してあるものをその都度出庫して用いた。結果(第177表)で注目されるのは、二重包装のものの孵化が普通包装のものに比べて1日あまり遅かったことである。これは、二重包装のものは、郵送中(速達便で送ったので、発送の翌日に明石に到着した)の暖房の影響を受けることが少なかったことを示すものである。おそく送ったものほど孵化日がおくれているのは、出庫日がおそいために外ならない。受取った蚕種を明石で冷蔵しておき、全部を同時に出庫して高温で催青したならば、催青日数が短縮するので(第150表)、孵化の遅速がこれほどはっきりとは確認できなかったであろうと思われる。
| 郵送 番号 |
包 装 | 孵化頭数(頭) | ||||
| 4月12日 | 4月13日 | 4月14日 | 4月15日 | 4月16日 | ||
| 1 | 普 通 二 重 |
2 0 |
50 1 |
481 226 |
7 178 |
0 16 |
| 2 | 普 通 二 重 |
0 0 |
45 4 |
478 424 |
2 37 |
0 1 |
| 3 | 普 通 二 重 |
5 0 |
86 4 |
365 427 |
9 86 |
0 5 |
| 4 | 普 通 二 重 |
4 0 |
40 0 |
476 370 |
8 111 |
2 17 |
| 5 | 普 通 二 重 |
25 0 |
62 0 |
445 342 |
5 260 |
0 6 |
| 6 | 普 通 二 重 |
12 0 |
23 0 |
449 177 |
4 296 |
0 13 |
| 7 | 普 通 二 重 |
0 0 |
12 0 |
517 58 |
6 431 |
0 25 |
| 8 | 普 通 二 重 |
1 0 |
65 0 |
492 5 |
3 416 |
1 59 |
堀口(216)は、輸送中の障害を考えて、どれぐらいの高温が何時間ぐらい続けば蚕種に害があるかを試験した。それによると、浸酸翌日から2.5℃に23日間冷蔵した青熟の即浸種を用い、鈴木ら(899,900,901)によって高温に対する抵抗力の最も強いと云われている催青4日目に、普通に考えられる範囲の種々な高温で処理したところ、対照区の孵化歩合が96.4%であったのに対して、試験区の孵化歩合は第178表の通りであった。
| 高温(℃) | 乾湿球の差(℃) | 接触時間(分) | 孵化歩合 |
| 50 | 10-11 | 15 45 |
約10%低下 孵化皆無 |
| 45 | 6.5-8 | 60 180 |
約10%低下 孵化歩合10%以下 |
| 41 | 4-6 | 120 | 約10%低下 |
| 38 | 5.5-8.5 | 240 360 |
6-7%低下 約10%低下 |
| 35 | 6-7 | 480 | 約10%低下 |
また、輸送用の円筒に蚕種を入れて地上5尺の炎天下におき、その影響を調べたところ、
38.5℃ 4時間で 孵化歩合の低下約5%
39.2℃ 6時間で 〃 約20%
であった。但し、温度は円筒の上部に設置した寒暖計によって測定した。
これらの試験において、高温に接触させた蚕種から孵化した蚕を飼育した結果によれは、41℃に接触させたものは孵化歩合の低いほど減蚕歩合(全令)が高かったが、40℃以下の温度に接触させたものでは、孵化歩合の低下と共に減蚕歩合の増加したものと、増加しなかったものとが相半ばしていた。
輸送中に、蚕種がこの程度の高温に接触する機会は少なくないと考えられるから、注意しなけれはならない。
飛行機でタト国へ蚕種を送る場合には、飛行機により、積載場所によって、高空を飛んでいる間は0℃近くまで下り、地上におりると40℃に近い亜熱帯の高温に接触することもあると云うから、これらのことを調べた上で、それに対応する荷造りの必要な場合がある。自動車も、構造によっては、排気管のそばに積込まれた蚕種が、その熱によって被害を受けた例があると云う。後に述べるように(XI1Bb)、60℃以下の温度に短時間接触して死んだ卵は、早斯死卵でありながら、催青しても潰れ卵にならず、孵化の間近かになって初めて気付くような場合もある。
人工孵化種および活性化した越年種は、輸送中に胚の発育が進み、受取った側でこれを勝手なときに冷蔵すると孵化を悪くすることがあるから、その点の連絡も必要である。冷蔵浸酸種の場合には白ハゼ卵防止の注意も大切である。
時差の関係で日付けが1日違うこと、途中の高温、高空を飛ぶための気圧の関係(Ⅵ3Dd)、それに、日本では、掃立てのときに全部孵化し切っているように、余裕をみて催青する習慣のあることなどが重なったためか、輸出した冷蔵浸酸種が予定の掃立て日よりも2-3日早く孵化して、大損害を受けたと云う苦情のきた実例もある。冷蔵浸酸種の孵化の揃い具合や、受取った蚕種の取扱いについての経験の浅い国への蚕種輸出には、このような点についての連絡や注意も必要である。
人工孵比種の荷造りには蚕種の呼吸も考えなければならないが、浸酸後2-3日の間は呼吸量が少なく、また段ボール箱などはガラス器とは違うから、呼吸量から計算した所要空気量そのままの容量の外装をする必要はない(Ⅵ3Ba)しかし、時期と場合とに応じて、内部に収める各蚕種容器の間には間隙を設け、空気の流通をはかり、蒸れを防ぐ必要がある。
自転車の荷台に蚕種や蟻蚕の入った容器をのせて運び、直射日光にさらすような不注意も避けなけれはならない。
産卵直後に郵送する程度の動揺は普通には孵化に影響がない(48)。
XI 蚕種の事故
保護、取扱いの過程で偶発した蚕種の事故には、その原因が正しく究明されず、従って蚕種保護の改善に役立つことなく葬られてしまうものが極めて多い。事故原因の究明が正しく行なわれ難いのは次ぎのような理由によるものである。
1)手近かな処に原因を求めて処理してしまう。例えば、越年種に早期死卵が多かったときに、たまたま夏期が高温であると、原因をそれに求めがちであるが、後に述べるように、早期死卵と考えられているものにも原因が極めて多いから、簡単に判断することはできない。
2)事故が公表されない。研究所や試験場とは違い、蚕種業者で起った事故は、営業政策上、ありのまま公表されず、内部で処理されてしまうことが多い。
3)事故に気付くのがおそい。外見でわかるような形態変化を伴なって蚕種が即死するような場合を除けば、一般には、原因の作用したときと事故に気付いたときの間にはかなりの日時の経過しているのが普通で、越年種の場合には特にこのような事故が多い。気に留めないで過こしたことをあとで思出そうとしても、正確な記憶のないのは当然で、強いて思出そうとすると記憶違いの混入するおそれもある。記憶違い、記録の誤まりなどは原因検討の妨げになる。
4)原因の隠されていることがある。担当者には、ありのままを報告しにくい事情のある場合が多い。試験場などへ事故原因についての相談のある場合にも、思い当ることはないがと云うだけで、責任逃れのために答弁資料を集めているのではないかと考えられるようなことが少なくない。
これに関連したことであるが、そんな筈はないと考えることも原因究明の妨げになる。
例えば、東京杉並郵便局から朝の9時受付けで、日野市(1955年当時は日野町)へ蚕種を送る試験を繰返えしたことがある。輸送の途中で高温障害を受けることがあるか否かを確めるために行なったもので、前記の明石への郵送も同じ試験の一部である。11回送ったうちの4回は6時間ぐらいで到着したが、あとの7回は22時間以上もかかって配達された。郵便局に問合わせても、速達であるからその日のうちに到着する筈で、途中で1晩泊るような筈はないと云うだけで、納得のゆく説明は得られなかった。しかし、何処かで1夜泊ったことは間違いないのである。同じ東京-日野間でも、日野から東京へ送ったものは、11回が全部8-9時間で到着した。
このような情況の下で蚕種に起こった原因を追究するためには、あらゆることを疑ってみて、原因を想定した実験を繰返えし、その結果から推定しなければならないことが多い。
河北・河田(361)は、永年に亘って蚕種の事故を調査した結果を第179表のようにまとめている。この原因の判定も上記のような意味で疑ってみなければならないが、一つの参考資料として示す。
| 事 故 の 原 因 | 原因別歩合 | |
| 歩合(%) | 原因分類 | |
| 1.品種的に多糸量で原種虚弱性の後作用 2.発蛾不一致、産繭抑制による不受精卵多量 |
0.7 0.7 |
計1.4% 卵素質不良 |
| 3.産卵当時の累積呼吸障害付与の形跡 4.春製越年種、夏期高温(80゚F%以上)の生理的消耗 5.春製蚕種、秋期漸降保護不完全または暖冬異変 6.洗落作業による物理的または温度衝撃 7.冬期冷蔵保護中長距離輸送移動の不完全 8.冷蔵以降最長期前まで胚子発育階梯別不適温 9.冷蔵末期過度の低温、活性化不斉遅延 10.人工孵化法(冷蔵時期、日数、温度、浸酸法)失敗 11.冷蔵保護温度激変に伴う白死卵発現 |
- - 1.4 1.4 0.7 11.2 4.9 5.6 7.0 |
計32.2% 蚕種保護 浸酸取扱不良 |
| 12.輸送障害(高温、蒸熱、遠距離、到着後解荷遅延) 13.催青着手時一定期間の予備催青不完全 14.催青温湿度環境の不備(主として低温低湿) 15.掃立日決定の慎重さを欠く(2夜包か2番掃か) 16.催青設備不完全(狭隘、挿換、光線、酸素不足) 17.眼点期以後暗室発蟻斉一化法の不徹底 18.催青卵および蟻蚕抑制方法、設備等の不備 19.掃立当日の取扱不良(当日の長距離運搬または明光線不足) |
9.8 1.4 15.4 1.4 6.3 18.1 11.9 1.4 |
計65.7% 催青取扱不良 |
| 20.卵数調査法の誤と相互確認連絡不良(小事も大事となる) | 0.7 | 計0.7% 連絡不良 |
蚕種の事故については、冷蔵浸酸種の冷蔵時期を誤まると白ハゼ卵が出る、と云うように要因になる事項を取上げて説明するのが普通であるが、白ハゼ卵の原因にも種々あって、個々にかぞえ上けれはきりがないから、ここでは、これとは反対に事故が発生したときに、その原因を考えるのに参考になる事柄について記述する。
1 事故蚕種の形態と原因
事故原因を検索する便宜上、蚕種の事故形態を13に分類して説明する。外観的には同じような死卵でも、長期の高温接触による死卵と長期冷蔵による死卵とは卵内容の変化が異なる筈であるが、これを識別し得る精密検査法は現在殆どあるいは全く未開発の状態である。
なお、薬品や放射線などによる蚕種の事故や、親に特殊な薬剤(例えばapholate)を与えた場合の死卵などは、元来蚕種に近付けるべきものではない要因の影響であるから、これらには言及せず、蚕種保謹上問題になることの多い要因だけについて述べる。
A 無着色卵
越年種にみられる無着色卵は普通に不受精卵と云われている。大部分は不受精卵と考えられるが、受精後、漿膜の形成される前に死んだ卵は、外観からも組織学的検査によっても、不受精卵または不完全単為発生卵と区別することがむずかしい。これらを区別せずに、不受精卵と呼んでいる。
従って原因は極めて広汎で、あらゆることが関係しているから、少なくとも上蔟以後の保護環境にまでさかのぼって検討しなければらない(Ⅲ3)。蛾の抑制(Ⅳ4)、交尾(Ⅳ3)および産卵(Ⅳ5)の管理などの採種に関する条件が直接に影響をおよぼすことは云うまでもないが、同じ母蛾の産卵でも初期よりは末期に多い(65)。遺伝的な欠陥によるものもある(Ⅰ2Ba、Ⅰ2Cb、Ⅰ3C)。
これらの卵は早期に潰れるのが普通であるが、保護環境によって異なり、一般的に云えば、春採りには潰れるものが多く、晩秋採りでは潰れないで残っていることが多い。
産卵の翌朝既に潰れているような卵は単なる不受精卵ではなく、卵殻形成に異常のあるもので、蛹を長く冷蔵した場合などに発現し易い(Ⅲ4A)。
交尾の欠陥や生殖器の異常による不受精卵は一般にかたまって産付けられており、蛹や蛾の抑制、雄蛾の使用過度などによる場合には、不受精卵と受精卵とが入り混って産み付けられている。
即時浸酸種の場合には、無着色の早期死卵と不受精卵との識別が更に困難である。浸酸による潰れ卵と不受精卵とを厳密に識別するには組織学的に漿膜形成の有無や胚組織の形態を調べなければ不可能であり、この方法によってもなお識別困難なことが多い。普通には無着色潰れ卵を不受精卵とみなして対受精卵孵化歩合を求めているが、即時浸酸種の場合には、無浸酸の対照区を設け、その不受精卵歩合によって更正しなけれは正しい対受精卵歩合は求められない。
真の不受精卵には分裂核の増殖が起こらない。単為発生が行なわれても、分裂核の増殖は受精卵におけるよりも遙かにおくれるから、産卵後6-10時間目頃(25℃)に卵を圧し潰して、内容を0.5%ぐらいのメチレンブリュー液(0.8%食塩水に溶解)で染色すると、分裂核があれば青く染まるので、産卵後間のない時期ならば、生物学的にはある程度不受精卵の識別ができるが、実用的ではない。産卵後25℃で20時間以上になると、受精卵には卵黄分割(Ⅵ2Ba)が始まっているから、生理食塩水中で生体解剖を行なうと、卵黄細胞の有無によっても受精卵か否かがわかる。この時期になれば受精卵には胚帯が形成されているから、Ⅶ2Aaで述べた方法によって外部から胚を透視することもできる。産卵後日のたった卵にはこれらの方法を適用できない。
B 着色死卵
a 催青着手前の越年種にみられるもの
これには原因の異なるものが種々含まれている。不受精卵の着色した所謂赤死卵や単為発生卵の潰れたものなども、早期死卵としてこの中にかぞえられていることが多い。
有賀・吉武(65)は、日122号の異常卵を、1)不受精卵(着色しないもの)、2)漿膜色素異常卵(非常に着色のうすい卵、漿膜に色素の局在している卵および白ハゼ卵など)、3)つぶれ卵、4)褐色卵(褐色を呈し、従来早期褐色死卵と考えられたもので、不受精卵の着色したものが多い)、5)濃色卵(後に述べる)、6)再出卵、7)催青死卵 に分類し、産卵後、同じ卵について継続調査を行ない、その変化を調べたが(第180表)、それによると、日のたつに連れて、不受精卵が潰れ卵や褐色卵に変化することが窺われる。
| 異常卵 | 調査時期 | |||
| 6月 | 8月 | 11月 | 3月 | |
産下卵 再出卵 漿膜色素異常卵 不受精卵 つぶれ卵(不着色) つぶれ卵(着色) 濃色卵 褐色卵 催青死卵* |
粒 351.1 0.0 1.9 14.5 0.1 0.0 0.0 0.0 0.0 |
粒 0.5 0.6 1.1 5.7 5.4 0.3 4.2 0.0 |
粒 0.8 0.0 0.0 5.7 14.8 1.8 0.2 0.0 |
粒 0.8 0.0 0.0 5.2 15.8 1.8 0.2 7.8 |
| 異常卵合計 | 16.5 | 17.8 | 23.3 | 31.6 |
褐色潰れ卵が不受精卵の着色によって生ずることは、佐藤ら(828)の調査によって極めて明瞭である。7月11-18日に28、25および22℃の温度において産卵させた日122号の卵を、9月20日までそれぞれの温度で保護した後、自然温度(新庄)に移し、1月下旬に調べた結果によれば、予め不受精卵を除去しておいた区には褐色潰卵は殆ど発現しなかった(第181表)。
| 保護 温度 |
A 不受精卵 | B 越年卵(不受精卵を除去せず) | C 越年卵(不受精卵を除去) | |||||
| 着色卵 | 無着色卵 | 褐色潰れ 卵 |
早期死卵 および 無着色卵 |
正常卵 | 褐色 潰れ卵 |
早期死卵 | 正常卵 | |
| ℃ 28 25 22 |
% 23 96 95 |
% 76 3 4 |
% 13 2 4 |
% 26 0 3 |
% 60 97 92 |
% 0 0 0 |
% 0 0 1 |
% 99 99 98 |
この場合、Aの28℃区において着色卵の少ないことは、高温によってオキシキヌレニンの自働酸化が促進されて不受精卵の着色が起こると云う有賀らの考えと相反するようにみえるが、そうではなく、28℃区においては、早く潰れ卵になって乾くために、最初の著しい着色卵の増加が途中で停止したのである。
白ハゼ卵のような漿膜異常を除けば、外形で区別のできる早期死卵は潰れ卵であると云ってよいが、このような着色早期死卵について考えてみると、第180表においては、不受精卵の着色したものが潰れたのと漿膜着色後の潰れ卵とが区別されていない。しかし、産卵後の経過に連れての不受精卵(無着色)および漿膜色素異常卵の減少と不着色潰れ卵および褐色卵の増加とを差引きすると、3月現在の着色潰れ卵の中の約1/3(供試部数の約1.5%)が漿膜着色後に死卵になったものであり、残り約2/3は不受精卵の着色によるものと考えられる。第181表においては(Cの場合)、漿膜着色後の死卵は最高1%であった。
これらは1月または3月における調査であるが、この頃に不良卵を全部除去して外観正常な蚕種だけを冷蔵しておくと、その後、春蚕催青のときまでの間に新たに潰れ卵の発生することは殆どない。
即ち、遺伝的に欠陥がなく、保護環境にも特別な異常がなけれは、正常に受精し、着色した越年蚕種が催青着手までの間に早期死卵の形態を現わすことは比較的少ないのが普通であるから、もし有色潰れ卵が非常に多いような場合には、不受精卵の着色とよく区別した上で、慎重に原因を追究する必要がある。不受精卵の着色との識別は、先きに述べたように厳密に云えば不可能な場合もあるが、実用的には漿膜形成の有無を基準にし差支えない。潰れ卵は温湯に浸して膨潤させて解剖する。
赤死卵と云われているものには褐色卵およびこれよりも一層赤味のある卵が含まれている。不受精卵(受精後間もない死卵を含めて)が着色し、湿度の関係で潰れずに残り、半熟鶏卵のように表層部が固まり、内部からの水分の蒸散をさえぎっているもので、色素は漿膜ではなく卵黄内で形成されている。赤死卵は洗落しの際の比重選や風選によって除去することがむずかしい。産卵後1ヵ月余りの間の保護湿度を60%以下にして、不受精卵を早く潰してしまえばその発現を少なくすることができるが、不受精卵を作っておいて、これを除去する方法に苦心するよりも、不受精卵を出さないようにするのが正しい対策であろう。またこれを早期死卵であると考えて、夏期の保護条件との関係が検討されたこともあるが、死卵ではない。夏期の高温期間に増加することは確かであるが、不受精卵が変化して生ずるもので、保護温度が比較的高く、多湿な場合に発現が多い。
単為発生卵には種々な発育程度のものがあり(Ⅵ2A)、漿膜の着色するまで発育したものが潰れると正常発生の着色死卵との識別ができなくなる。潰れる前の単為発生卵は、前極部付近だけが着色したり、着色した漿膜細胞が大形で配列が異常である、なでの特徴によって識別できる場合が多い。
濃色卵と云うのは、正常卵よりも濃色で、漿膜面に油を塗ったような感じのする卵である。孵化せず、死卵になる。高梨・松尾(956)によれば、春採りの日8号を12月に調査した処、180蛾中の56蛾にこの卵を認めたが、発現は少なく、総卵数(56蛾の)32,102粒中に229粒(0.7%)であった。濃色卵の発現の特異な点は、これが必らず不受精卵に接して発現していることで、1粒の濃色卵が1粒の不受精卵に接して発現していた場合が125例、2粒以上の不受精卵に接していた場合が25例、褐色死卵を隔てて不受精卵に接していた場合が19例で、不受精卵に接していなかった場合は1例もなかったと云う。有賀・吉武(65)は、濃色卵が不受精卵あるいは潰れ卵に近接して存在することから、これらの卵の胆汁が健康卵に付着して濃色卵を発現させるのではないかと考えているが、普通の潰れ卵は機械的に圧し潰したものではないから、どのようにして卵汁が飛出して隣接した卵に付着するのか不明である。有賀らは、人工的に卵汁を塗ると濃色卵ができること、第二白卵に塗ると濃色卵になるが、第一白卵に塗っても濃色卵にならないことなどから、不受精卵の着色と同様に、オキシキヌレニンの自働酸化によって生ずるものと考えている。濃色卵の胚の状態については記載されていない。
卵汁の付着した卵が死卵になることについては関戸(834)の報告がある。
卵のタト部からはみえないが、解剖したり、切片にしてみると、越冬期の胚のある部分が内方へ折れ込んだように屈折し、その前後の部分は正常に卵表に沿って位置している場合が縷々認められる。和田(1131)は最長期(丙B)以後の卵にはこのような胚のみられないことから、これが死卵の原因になることはなかろうと考えている。普通の場合には、おそらく回復して正常に孵化するものと思われるが、胚の内方へ折れ込んだ部分が羊膜を伴なっていないことは、胚の伸長が始まったときに羊膜の伸長が伴なわず、その抵抗によって胚が折れ曲ったと云う感じであるから、長期間冷蔵を続けて卵の活力が低下すると、回復し得ないで、戸谷(1063-1065,1067)のみているような屈曲あるいは捻転胚に発達するのかも知れない。
b 越年種の催青を始めてから発現するもの
催青を始めると、これまで外見では識別のできなかった死卵が特徴を現わし、また活力は低下しても低温のため辛うじて生きていた卵が、発育を続け得ずに死卵になることもあって、種々な死卵が発見されるが、原因は単一ではない。
夏期保護が適当でなかったための死卵も、冷蔵が適切でなかったための死卵も、余程の場合でなければ、そのとき直に死ぬことはなく、次第に消耗し、催青を始めてからの死卵になるものが多いが、被害の程度によって、早期死卵になるものから催青死卵になるものまで様々であるから、卵の死んだ時期の遅速よって被害を受けた時期が推定できるとは限らない。例えば、第104表において、催青第1日から第12日までの間の各期に低温接触を行なった揚合の死卵は、接触時期に関係なく殆ど総べて催青死卵であった。
また、早期死卵でありながら催青中に潰れず、正常のものが点育してから初めて気付く場合もある。適当な高温に短時間接触して死んだ卵や、洗落しの後、濡れたままのバラ種を長時間堆積しておいたために蒸れた卵などにこのようなことがみられる。
1例を挙げると、一蛾採りの越年蚕種を1月下旬に郵送したものを、4月下旬から催青した処、第30図に示すように、1蛾の卵の中に局部的に死卵の発現した場合が発見された。

第30図 郵途中に高温に接触した死卵の1例(高見)(925)
黒ぬり:早期死卵、二重線:漿膜に小円形の破れのあった
早期死卵、太線:催青死卵、細線:正常に孵化した卵
死卵の分布状態からみて、死卵を発現させた要因は図の右側の方向から働き、強い作用を受けた部分の卵は早期死卵、弱い作用を受けた部分の卵は催青死卵になったと考えられるものであった。この標品を入手したのは催青後2ヵ月経過した7月であったが、早期死卵には、潰れたものの外に、水引きの強い程度の凹み方で残っているものが多かった。解剖してみると胚は甲から丙Bぐらいまでの間で死んでいた。
このようにはっきりした形態および分布を示す卵の死に方は、薬品その他の影響としては考えられないので、種々な実験を重ねた結果、55℃の湯に1分、または60℃の湯に5秒間ぐらい浸して殺した卵は長期間に亘って潰れないことがわかった。このような卵を固定せずに生理食塩水中で解剖し、メチレンブリューで染色すると、胚、卵黄核および蚤自性卵黄顆粒の染まり具合、脂肪性卵黄顆粒の凝集状態などに特徴のある種々な変化がみられた(932)。固定して染色すると、全体一様に染まるからデリケートな変化がわからない。そこで、融点のわかっているパラフィンを塗布した紙を蚕種の間に挟んで郵送箱に収め、外部から高温を作用させて、死卵の発生とパラフィンの融解状態との関係を調べ、更にⅩ3で述べた郵送実験を経て、問題の死卵が郵送中の高温接触によるものであることを確認した。
洗落し時期および洗尽し後の取扱いが悪いために胚が不時発生を始めた場合、および軟性卵で活性化比が早かった場合などにも種々な時期の早期死卵が発生する。
このように早期死卵の原因は極めて多いから、卵の外観および解剖による調査ばかりでなく、再出卵、点催青死卵、不受精卵、孵化の斉否および早晩などとの関連にも注意し、あらゆる方面から検討しなければ原因を究明することはむずかしい。
c 人工孵化種にみられるもの
人工孵化性においては、上に述べた越年種の場合と同じ原因によるものの外に 浸酸および冷蔵による死卵が発生するから、それらの項で述べた問題をも考慮に入れて検討しなけれはならない。
C 淡色卵
漿膜色素は主として3・ヒドロキシキヌレニンに由来するが、その母体から卵内への移行および卵内での消費は越年性の不十分な卵が多い(Ⅶ1A)。越年性の不十分な卵は再出卵や孵化不良の原因になりがちである。
従って、卵色の淡いことが間接に孵化不良の原因に結び付く場合があるが、ただ間接的な意味ばかりではなく、吉川(370,372)は、第一白卵種の5令幼虫に正常卵色系統の蚕の卵巣を移植して、白卵に着色を起こさせると、その卵の孵化歩合が向上し、卵色の濃いものほど孵化歩合の高いことを認め、着色現象そのものに何らか孵化をよくする作用のあるらしいことを報告した。その機構は明かでない。
実用問題としては、三谷(540)が春採りの日115号および支108号を用いて試験を行なった。産卵後1ヵ月たった蚕種から死卵を除き、卵色の濃淡によって5階級に分け、それぞれ7月1日から8月31日までは次ぎの温度におき、
高温区 85゚F(29.4℃)目標
常温区 75゚F(23.9℃) 〃
低温区 70゚F(21.1℃) 〃
9月1日以後は何れも同じ冷涼な蚕種庫に保護し、1月から5℃、4月20日出庫、中間温度3日を径て77゚F(25℃)で催青したところ、高温区は両品種とも孵化が悪かったが、特に卵色の淡いものの孵化が劣り、卵色の階級と孵化との関係は支108号において極めて明瞭であった。常温区および低温区においては、支108号は何れの卵色のものも孵化がよかったが、日115号は、高温区に比べて一般に孵化がよかったとは云え、矢張り、淡色および最淡色卵として区分したものの孵化が劣った。
これによると、卵色の淡い卵の虚弱なことが明かであるが、保護温度および品種によって、その影響の現われ方が相違する。孵化しなかった卵の形態や胚の状態については記載されていない淡色卵の原因にも言及されていないので、対策は不明であるが、この場合には、3・ヒドロキシキヌレニンの移行および消費に関係のある条件と共に、遺伝的な性質および初期産卵と末期産卵とによる卵色の違い(Ⅶ1A)も関係しているように考えられる。末期産卵は卵が小さく種々な点で劣ることが知られている(Ⅱ2Ac)。
D 白ハゼ卵
白ハゼ卵は中間温度を通さずに、蚕種を高温の場所から急に低温の場所に移した場合に発生し易く、特に冷蔵浸酸種の再冷蔵によって起こり易い(Ⅷ2Ac、Ⅷ3C)。白ハゼと云うのは、漿膜が破れて1ヵ所に凝集した異常の総称であるから、原因は様々で(658)、その発現の程度も一定しない(581)。
遺伝的な白ハゼ卵として報告されているものも多いが(41,44,45,674,1107,1109)、この中には冷浸種の再冷蔵による場合とは現れ方のかなり異なったものが含まれているようである。遺伝的とは云っても、その発現には保護、取扱いの影響が大きい。支127号および銀白(1028)の白ハゼ卵は夏期の高温保護期聞か長いと多発する。
白ハゼは漿膜が破れて凝集するものであるが、漿膜は風船羊羹のゴム膜のように緊張しているものではないから、その一部分を傷つけても、傷口が大きく開いて、傷とは反対の側に向って膜が収縮するとは限らない筈である。実際にも、産卵後40-50時間目頃に強い浸酸を施すと、漿膜には明かな亀裂が生じるが、それが拡がらず、寧ろ、破れた端の細胞が集合、収縮して黒色の斑点を形成し、破れを塞ぎ、白ハゼに発展しない。沓掛(473)も再冷蔵を行なわない冷蔵浸酸種について同様の現象を観察している。
これに対し、冷蔵浸酸用に蚕種を冷蔵して20曰ぐらいたつと、漿膜面に黒い斑点が現われるが(15)、この黒斑は細胞の収縮した部分であり、これが周囲の細胞を引き寄せている。黒斑部の細胞がどのような機構によって収縮し、周囲の細胞を引き寄せるのかは不明であるが、このような張力が働けば、損傷の生じた場合に、破れが拡大して白ハゼに発展することは十分に考えられる。
高温で保護しておいた卵を急に低温に移し、白ハゼ形成の過程を観察すると(473,924)、まず卵内から液体が漿膜面に溶出してくる(XI1E)。これは急激な温度変化によって漿膜に何らかの損傷の生じたことを示しているが、低温においたままではそれ以上に発達しないのが普通である。このような卵を高温に移すと(再冷蔵した冷蔵浸酸種について云えば、その出庫)、初めて漿膜に亀裂が生じ、漿膜細胞の収縮、凝集が起こって白ハゼ卵に発達する。
白ハゼ卵の発現に種々な生理的要因の関係していることは勿論であるが(Ⅷ3C)、機械的には、第一段の漿膜の損傷に第二段の漿膜細胞の収縮、凝集が伴なわなければ、典型的な白ハゼには発達し得ないもののようてあり、細胞の収縮、凝集が高温に戻してから促進されることを考えると、この生理的な要因は第二段の変化の進行に関係しているように思われる。
蚕卵をアセトンガスで処理すると漿膜が破れて収縮し、白ハゼ類似の形態を現わすが、アセトン中に浸漬すると却ってこの発現が少ない(924)のは、障害が強過ぎて、第二段の変化に必要な細胞の生理機能が損なわれるためではなかろうか。クロロホルムやエーテルで処理すると、卵液の滲出はアセトン処理の場合以上に著しいが、白ハゼ類似の形態に発達しない。第30図において、孵化卵と死卵との分布の境界部に、漿膜に小さな円形の破れを現わした死卵が若干みられたが、この破れが円形を呈したことは、損傷卵の漿膜が周囲から引かれて発現したことを示しているにも拘らず、典型的な白ハゼに発達しなかったのは、漿膜の受けた障害が大きく、第二段の変化の進行に必要な生理作用が停止したためではなかろうか。
荒木(59)が、白ハゼ卵は卵を即死させるような条件の下では発現せず、徐々に発育しながら致死するときに起こると考えているのも同様なことを指すもので、白ハゼの発現に卵の生理状態の影響の大きいのもこのためであろう。
卵殻を除去し、漿膜に包まれた卵内容全体を卵外で培養しながら観察すると、産卵後日数のたっていない卵においては、漿膜表面のクチクラ(Ⅵ2Ba)が漿膜に密着していて離れにくいが、この頃の卵の漿膜は亀裂を生じても、それが拡大して膜の凝集することが少ない。これに対して、産卵後2日の卵を冷蔵し、1ヵ月ぐらいたって、再冷蔵による白ハゼ卵の発生し易い時期(1150)になると、漿膜とクチクラとが離れ易くなる。また、白ハゼ卵を切片にしてみると、漿膜の凝集した部分においては、漿膜とクチクラとの離れが特に顕著である。
これらのことを考見合わせると、漿膜とクチクラの付着状態もまた白ハゼ発現の第二段の変化に関係があり、漿膜とクチクラとが密着していると凝集が起こりにくいのではないかと云う推測もできる。
しかし、卵の内容を培養して、漿膜とクチクラとの離れる状態を観察すると、離れた漿膜細胞の一部がクチクラに付着したまま各所で細く引きのばされているのをみることがある。特に、長く冷蔵した卵なでにおいてこれが多くみられるから、急激な温度変化によってクチクラから引き離されると、漿膜細胞が損傷して白ハゼ発現の第一段の条件が成立することも考えられる。
白ハゼ発現の機構には、このように、物理的な傷害と細胞収縮の問題とが結び付いていて興味があるが、現在の処では現象的な研究が行なわれているに過ぎない。
白ハゼ卵を低倍率の顕微鏡で観察すると、白くなった部分の表面に、褪色した漿膜が残っているようにみえるが、これは凝集する以前に付着していた漿膜細胞のプリントがクチクラの面に残っているのであって、漿膜ではない。
白ハゼ卵の発現には種々な程度のものがあるが、胚発育の異常も様々で、孵化するものもある。
E 卵液の滲出
卵の内容成が漿膜を通し、その外面へにじみ出てくる現象で、卵の後極部において卵殻と卵内容とが離れ、その間隙に液の溜ったものが最も普通の発現状態である。
卵液は蛋白質を含んだ濃厚な液で、正常な卵においては漿膜を通過することはないが、漿膜が損傷すると、外観では漿膜の破れの認められない場合にも滲出が起こる。白ハゼ発現の第一段の変化として現われる場合については前項で述べた。
洗落しの際の揉み方が強かったり、乾燥の際の麦とのぶつかり合いが激しかったりした場合(906)、落下衝撃を受けた場合(939)、出庫直後の浸酸(451)などにも発現する。漿膜細胞のいためられるような条件は総て原因になるから、高温保護の長かった卵にもみられる。これが白ハゼに発達するか否かは、第二段の細胞収縮を起こす条件の有無によってきまるから、極く軽微な白ハゼ卵と考えてもよい。
卵液滲出卵の孵化歩合は障害の程度によって異なる。孵化したものの飼育成績は、高橋(906)の調査した洗落しによる滲出卵においては、正常卵と異らなかった。
卵液の滲出は物理的な損傷の外、クロロホルムやエーテルのような脂肪溶解性の薬剤によっても発現する。カルノア液で蚕卵を固定すると、固定の悪かった場合に、漿膜面に白い沈澱ができて白ハゼ類似の外観を呈することがあるが(667)、これはクロロホルムの作用によって漿膜細胞の構造が破壊され、卵液が滲出し、アルコールによって凝固するのであって、白くは見えるが漿膜が破れて一方に寄るのではない。このカルノア液による白色沈澱が日本種の卵に生じ易く、支那種の卵には少ないと云われ、白ハゼ卵の発現と相似た関係を示すのは、日ハゼの起こり易い日本種の卵は漿膜細胞の損傷を生じ易いためかも知れない。
F 卵黄膜の剥離
Eの場合には卵殻と卵内容との間に卵液が溜るのであるが、液が溜らずに卵殻と内容との離れる場合がある。小さな三日月形あるいは三角形の間隙が普通で、液の代りに空気が入っているので、白くみえる。
蚕卵には、第23図に示したように、卵殻の直下に卵黄膜がある。この膜は、生のまま卵殻をはがすと卵殻の内面に付着して取れ、出殻においてもその内面に付着して残っている(387a,983)。
剥離卵は、卵殻とこの卵黄膜とが部分的に離れ、そこに空気の入ったものが大部分(おそらく全部)であって、洗落しのときの衝撃や揉み方が強かった場合に起こるが、白ハゼ卵や滲出卵のような漿膜細胞の損傷がないため、卵液は滲出しない。
空気の入る機構は明かでないが、蚕卵をアセトンに入れて内容を膨潤させると(926a)、卵の腹面正中線または楕円の長軸に沿って縦に卵殻の亀裂するものの多い中で、処々に微小な点状の内容噴出を起こす卵のあることからみると、卵殻には、普通の状態では気付かない微小な欠陥や弱点があり、このような部分に損傷が生じるのではないかと思われる。
しかし、剥離の生し易いのが第23図の右下方のような端部であることを考えると、卵の部分によって、形態上、機械的衝撃の受け方が異なり、これと卵殻の弱点との組合わせによって、発現部位がきまるのかも知れない。
G 点催青死卵
点催青死卵は、胚発育の比較的後期にうけた影響によって起こるもののように考えられていることがあるが、実際には、あらゆる時期の影響がここに集約していると云うことができる。
弱い障害を受けている卵の胚や、発育機能の不十分な胚は発育の最後の段階で力がつき、漿膜を飲込むことはできても、卵殻を喰破り得ないで点催青死卵になることが多いから、活性化不十分で孵化機能の低い卵も、活性化が早過ぎて活力の低下した卵も共に点催青死卵の原因になる。堅い卵殻を喰破ることは最後の難関と云える。
鎌倉・金刺(304)は、冬期間の冷蔵温度が催青死卵の1原因であるとしているが、催青死卵の原因と云う立場からみれば、正に1原因に過ぎない。
第30図において、高温の影響を最も強く受けた部分の卵は即死し、影響の弱かった部分の卵は孵化し、その中間の部分に催青死卵の認められることは、同じ時期の同じ要因の作用によっても、その程度が相違すれば死卵になる時期の異なることを示している。
勿論、点催青卵になってからの悪条件によって死卵になることも多い。
DDTなとによって蚕卵が死ぬ場合に、多くのものが催青死卵になるのは、これらの薬品が神経毒であるために、神経器官が発達してから死ぬのであると説明されているが(543,1083)、蚕卵はDDTの溶媒になっている油剤だけの影響によっても死ぬのであるから、この場合にも、神経をおかされたため以外に、催青卵で力の尽きた卵が含まれているものと考えられる。
微粒子病に感染した卵も催青死卵になることが多い。
H 不孵化卵
これは他の卵が孵化してしまった後に、死卵の形態を現わさずに、催青着手当時と変らない外見のまま残っている卵で、活性化不十分な卵と考えられていることが多いが、この中には、全然活性化していないもの、不完全または畸形的な発育をしているもの(Ⅷ4)、死卵を不孵化卵と見誤まっているもの(XI1Bb)などが含まれているから、解剖して確かめる必要がある。
I 卵殼喰破りの異常
卵殻を僅かに齧っただけで孵化し得ないものは、催青中の取扱いが不良てあった場合の外、点催青死卵の場合と同様に前々からの障害の影響がここで現われたものも多い。
孵化時の物理的刺戟が原因になることも少なくないから、孵化調査の場合の取扱いや気流(640)などに注意が必要である。
正常の場合には、卵殻の前極部、腹側寄りを喰破って孵化するのであるが、ここを完全に喰破り得ないで卵内を転々し、腹側あるいは後極に近い部分などを齧っていることがある。原因は明かでないが、浸酸処理の不適当な場合などにもみられるようである。
J 再出卵
K 孵化不斉
孵化の揃わないのは活性化の不斉によることが多いが、その他、蚕種の取扱いが適当でなかった場合、障害を受けた場合などには、卵によって影響の受け方が異なるため、総て孵化不斉の原因になる。従って、例えば浸酸種においては、刺戟不足の場合にも過度の場合にも共に孵化が不斉になる。
孵化の際の温度、光線、湿度などの影響も著しい。
L 蟻蚕の畸形
Ⅷ4参照。
M 死蟻
掃立てのとき死蟻のあるのは、催青中および孵化後の取扱いの不適当なことが原因になるのは勿論であるが、蚕種保護が適当でなく、卵の活力の低下している場合に、孵化はしても蟻蚕の絶食生命時数が短かいため、2夜包み、3夜包みに耐えないで死ぬこと、卵の大小によって絶食生命時数の相違することなども原因になるから、あらゆる点に注意しなけれはならない。
蟻蚕が吐液しているのは毒物による中毒である。
2 病毒の経卵伝達
蚕種そのものの事故ではないが、蚕種を通して次代蚕に伝えられる蚕病は、影響の範囲が広いだけに、蚕種そのものの事故以上に重視しなければならない。卵を通して伝えられる蚕病として蚕種製造上最も重要なのは微粒子病であるが、これは法規にも定められている病気であり、著書も多いことであるから省略し、ここでは、最近、蚕種関係者の注目をひいているウイルス病の経卵伝達の問題について、蚕種の立場からの検討を加えたい。
家蚕において問題になっているウイルスの経卵伝達は主として細胞質型(中腸型)多角体病および伝染性(F型)軟化病についてであって、これらのウイルスが潜在状態で、母体から卵を通して次代蚕に伝えられると云うのである。
これは伝染とは違い、母体からウイルスを伝えられた筈の卵を磨砕し、蚕に接種しても感染は超こらず(942,1153,1179)、卵にも、孵化した蟻蚕にも異常は認められないが、飼育や取扱いが悪いと、潜在ウイルスが活性化して発病すると考えられている。従って、蚕種製造者にとっては、病毒を持っている卵を除去する方法がなく、しかも蚕作をみてから責任を問われるおそれのある点で、微粒子病の場合とは違った不安がある。
A ウイルス病の経卵伝達
家蚕のウイルス病が経卵伝達をすると云うことが初めて問題になったのは、次ぎの実験の結果からである(第182表)。
| 交 配* | 低温処理を 施した次代 5令蚕数 |
発病蚕数 | 病蚕内訳 | |||
| T | T>H | T<H | H | |||
0(H)♀×0(H)♂ 0(T)♀×0(T)♂ 1(T)♀×3(T)♂ 3(T)♀×3(T)♂ |
頭 17 82 18 66 |
頭 4 19 8 38 |
頭 0 4 1 2 |
頭 0 2 0 0 |
頭 1 0 0 8 |
頭 3 13 7 28 |
蚕の細胞質型多角体病ウイルスには、6角形の多角体を作るもの(H)と4角形の多角体を作るもの(T)とがあるが、一般に、HはTよりも広く分布している。5令期にこれらのウイルスを接種したところ、多数の病蚕が発生したが、生き残って化蛾したものを掛け合わせて採種し、次代蚕の5令起蚕に低温処理(5℃、24時間)を施してウイルス病の誘発を試験した。その結果によると、親の代にHを接種したものの次代にはHだけが発生し、Tを接種したものの次代には、Hの外に、Tが発生した。
親がウイルス病に罹っていると、その卵の外面にはウイルスの付着する機会が多く、従って、これを喰破って孵化した次代蚕がこれに感染したのではないかと云う疑いもあるが、次代蚕は加温即時浸酸種を掃立てたので、卵面は塩酸によってある程度消毒されているから、その後の汚染がなければ、この点の疑いは少ないと考えてよい。しかし、次代蚕の飼育を無菌状態で行なったのではないから、飼育中における自然感染が試験区によって相違し、偶然このような結果になったのかも知れないと云う疑いは残る。
そこで、飼育中の自然感染をできる限り均一にするために、次ぎのような方法で実験を繰返えした。
斑紋の異なる三つの大造系統、褐円(L)、虎斑(Ze)および黒縞(ps)を材料に用い、5令起蚕で、LにはHウイルス、ZeにはTウイルスを添食し、これらから採種した次代蚕をウイルスを添食しなかった(対照)psと混合して飼育した。混合しても3系統の蚕は斑紋によって識別できる。これらの5令起蚕に低温処理(5℃、24時間)を施した処、親に接種したのと同じ型のウイルス病が子の代に発生した(第183表)。
| 交 配* | 低温処理を 施した次代 5令蚕数 |
発病蚕数 | 病蚕の内訳 | |||
| T | T>H | T<H | H | |||
L(H)♀×L(H)♂ Ze(T)♀×Ze(T)♂ ps♀ × ps♂ |
頭 18 17 21 |
頭 6 11 10 |
頭 0 6 3 |
頭 0 5 0 |
頭 0 0 0 |
頭 6 0 7 |
ただ、対照蚕にTウイルスが非常に多く発生したことは、この実験条件の下においては、Tウイルスの自然感染の多かったことを示し、試験結果の解釈に若干の不安を抱かせる。
しかし、その後繰返えされた試験結果がこれと同様な経卵伝達の傾向を示しているから(64,67,1186)、このような方法によって実験を行なう限り、細胞質型多角体病ウイルスが潜在状態で母体から卵を通して次代蚕へ伝えられるのであろうと云う考えを否定することはできない。
B 経卵伝達の証明
ウイルスが卵によって伝えられる可能性のあることは上の実験によって示されたが、飼育中の感染を完全に防止して実験を行なわなければ、これを確証することはできない。
しかし、桑を与えて飼育するのでは無菌飼育が不可能なので、人工飼料を用いて無菌飼育を行なってみると、親にウイルスを接種しておいても、その子においてはウイルス病の誘発がみられない。蚕の体内に潜在状態で保有されているウイルスを活性化し、発病させる最も有効な手段は5令起蚕の低温処理であるが、無菌飼育の蚕には、低温処理ばかりでなく、誘発に有効であると云われている薬品やX線を併用しても誘発が起こらず、また、無菌飼育中に軟化病症状を呈して死ぬ蚕があっても、これには伝染性がない。このように無菌飼育をすると誘発の起らなくなるのは、細胞質型多角体病ばかりでなく、核型多角体病(膿病)についても同様である(460,461,503-508,1059)。しかし、このような無菌飼育の蚕にも、ウイルスを接種すると、多角体病ばかりでなく伝染性軟化病もよく感染する(18,20,508)。
これらの結果は、一見、経卵伝達は次ぎに述べる潜在感染ではないかと云う疑いを抱かせるが、桑で飼育しても、葉質や蚕の栄養状態が違えば低温処理による多角体病の誘発成績の相違することが知られているから(63)、桑とは著しく異なった飼料を与え、隔離した無菌状態で飼育した場合の誘発成績が相違しても不思議ではないとも考えられて、問題のきめ手にはならないのである。核型多角体病については、この点に関して、無菌飼育の蚕でも、これを空気中に暴露した後は、無菌のときと同じ飼料を与えていても、低温処理または薬品添食によって誘発の起こるようになることが報告されている(505,507)。
なお、柞蚕においては、無菌飼育の幼虫に、唯1頭ではあるが核型多角体病の発生した例が報告されている(1032)。
蟻蚕または稚蚕期にウイルスを接種しても、接種量が少なければ、ウイルスは体内であまり増殖せず、多角体も作らず、潜在感染の状態に止まっており、壮蚕期の誘発処理によって初めて発症することが知られているが(22,23,62,463-465)、無菌飼育が利用できなければ、このような潜在感染と経卵伝達とを明確に区別することは不可能能である。無菌飼育によって調べてみると、低温処理を受けた5令起蚕は一時的にウイルス感受性が高まり、感染し易くなるから(460)、低温処理によるウイルス病の誘発は、経卵伝達で伝えられた潜在ウイルスの活性化よりも、低温処理直後の感染によることが多いのではないかと云う疑いも残る(151)。
ウイルスの経卵伝達、潜在感染、誘発をめくる問題は、このように専門学者の間においてさえなお意見の一致をみていないのであって、蚕種の立場から割切った答を期待しても無理な現状である
C 蚕種の実際問題
蚕種に関する実際問題はどうかと云うと、横川ら(1187,1190,1191)の誘発実験によれば、即時浸酸については、産卵後24℃、20時間目に浸酸(比重1.075、液温46.1℃、5分間)を行なった場合には、産卵後15、25、30時間目の浸酸よりも多角体病(主として細胞質型であるが、核型をも含めて)の発生が少なく、冷蔵浸酸においては、出庫後24℃、3-5時間で浸酸したものには、その前後に浸酸したものよりも多角体病の発生が少なく、催青温度については、反転期を境にして温度を変化させた場合(25℃→30℃、30℃→25℃)または30℃平進の場合に比べて、25℃平進の場合に多角体病が少なく、6月20日産卵の越年種を1ヵ月間自然温度で保護した後、9月15日までを25℃で保護したものと32℃で保護したものとを比較すると、多角体病の発生は前者に少なく、活性化した越年種を2月中旬に、毎日午前10時から午後3時までの間、10日または20日間に亘って、20℃の高温に接触させたものから孵化した蚕には、このような高温接触を行なわなかったものに比べて多角休病の発生が多く、浸酸後の脱酸の不十分な場合を想定して、種々な濃度の塩酸を卵面に塗布した場合には、孵化歩合には差がなかったが、15%塩酸(普通の即時浸酸に用いる濃度)を10,000倍に稀釈したものを塗布(舌ざわりで僅かに酸味を感じる程度)した場合にも対照(水塗布)に比べて多角体病の発生が多かった。
樺沢ら(301)によれば、細胞質型多角体病の発生は、催青卵冷蔵(5℃)24時間までは違いがないが、72時間以上になると増加し、産卵後、即時浸酸までの保護温度については、25℃の場合に最も少なく、30℃がこれに次ぎ、20℃で最も多く、産卵から冷蔵浸酸のための冷蔵までの保護温度については、20℃および25℃に比べて30℃の場合に僅かに多く、冷浸種を14日間冷蔵した場合には即浸種を16日冷蔵した場合に比べて多く、産卵後、即時浸酸までの時間については、14-18時間に比べて21-22時間のものに多く、母蛾を5℃に1日または3日間冷蔵しても、即浸種における発病に影響がなかった。
引地(192-194)によれば、出庫直後に冷蔵浸酸を行なうと中腸型多角体病の発生が多く、加温と常温との即時浸酸を比べると前者に多かった。
これらの記載には、例えば、越年種の夏期保護温度が高かった場合に卵がどのような状態になっていたのか、産卵後14-18時間の即浸において発病が少なかったと云うが、その孵化歩合はどうであったかなど蚕種保護の立場からの問題点は多いが、注目されるのは、産卵後14-18時間目の即浸に少なかったと云う場合を除けば、総べて、蚕種保護の常識から考えて、取扱いの良かった場合には発病の少ないことである。
親の代に細胞質型ウイルスを接種した後、不良環境を作用させた実験によれば、稚蚕期の軟葉給与、壮蚕の硬葉(20-25葉位)給与、起蚕絶食、高温飼育(1-2令32℃、3令30℃、4-5令30℃)などは病蚕の発生を多くするが(1192)、緑蚕上蔟、種繭冷蔵、3夜包みなどは特に病蚕の発生を高めなかった。
5令起蚕に4角形細胞質型多角体病ウイルスを添食した後、緑蚕上蔟、種繭冷蔵、次代蟻蚕の3夜包みなどの不良環境を作用させた結果によると、母蛾に多角体の形成されていなかった場合にも(第184表)、多角体の形成されていた場合にも(第185表)、共に親の代に接種したと同じ型の多角体病が次代蚕に発生することは極めて少なかった。
| 不良環境の種類 | 1-2令 用桑 |
掃立~結繭の 病蚕歩合(%) |
種類別細胞質型多角体病蚕歩合(%) | |||
| T | T>H | T<H | H | |||
| 緑蚕上蔟 | 軟 葉 普通葉 |
34 56 |
0 0 |
0 0 |
0 0 |
34 52 |
| 種繭冷蔵 | 軟 葉 普通葉 |
10 74 |
0 2 |
0 0 |
0 0 |
10 68 |
| 催青卵冷蔵 | 軟 葉 普通葉 |
54 58 |
8 2 |
0 0 |
0 0 |
44 56 |
| 3夜包み | 軟 葉 普通葉 |
34 44 |
2 4 |
0 0 |
0 2 |
30 38 |
| 対 照 | 軟 葉 普通葉 |
26 50 |
0 0 |
0 0 |
2 0 |
24 50 |
第184表の場合には、添食したと同じ型の多角体が発見されなかったのであるから、あるいは感染しなかった蟻であるかも知れないので、その次代蚕において同じ型のウイルス病の発生が少なくても不思議はな.いが、第185表の場合には、多角体の形成を確認した感染蛾の次代蚕であるにも拘らず、添食したと同じ型の病蚕の発生は第184表の場合よりも少ないぐらいである。
| 不良環境の種類 | 1-2令 用桑 |
掃立~結繭の 病蚕歩合(%) |
種類別細胞質型多角体病蚕歩合(%) | |||
| T | T>H | T<H | H | |||
| 緑蚕上蔟 | 軟 葉 普通葉 |
62 76 |
0 8 |
0 0 |
0 0 |
60 68 |
| 種繭冷蔵 | 軟 葉 普通葉 |
64 48 |
0 0 |
0 0 |
0 0 |
62 48 |
| 催青卵冷蔵 | 軟 葉 普通葉 |
56 86 |
0 0 |
0 0 |
0 0 |
52 82 |
| 3夜包み | 軟 葉 普通葉 |
32 52 |
2 0 |
0 0 |
0 2 |
30 50 |
| 対 照 | 軟 葉 普通葉 |
26 50 |
0 0 |
0 0 |
0 0 |
26 50 |
これは、これらの不良環境には病気を誘発する作用のなかったことを示すだけで、経卵伝達を否定する結果ではないが、両表を通じて、添食しなかった筈の6角形多角体病の発生の著しいのに比べると、経卵伝達による発病の占める比率は極めて小さいもののように考えられる。
この自然発生の6角形多角体病が飼育中の感染によるものとすれば、経卵伝達による発病(無菌飼育ではないから、表に出ている4角形にも自然感染によるものが含まれている筈である)はこれに比べて極めて少ないと云えるし、もしまたこの6角形が、自然に感染し、自然に経卵伝達した潜在ウイルスによる発病を合んでいるものとすれば、経卵伝達は蚕には極めて普通であって、経卵伝達による潜在ウイルスを持たない蚕は寧ろ珍らしいと云うことにもなる。
母蛾のウイルス病症と次代蚕の発病との間に関係のないことは、細胞質型多角体病ばかりではなく、核型多角体病についても報告されている(19a)。
D 蚕種の立場からの対策
このように、ウイルスの経卵伝達は実際問題としても明確とは云い得ない上に、言葉としては新らしくても、新らしい病気が侵入して来たのではなく、事実についての解釈の論議であるから、特別に心配をすることはない。しかし、少なくとも経卵伝達と解釈されるような事実のあることには注意する必要がある。蚕種の立場から重要なことは、事実だけであって論議ではない。
従って、対策の第一は、原蚕を健康に育てることと、蚕種の汚染を防ぐこととである。蚕を健康に育てることの必要は云う迄もなないが、蚕種の汚染防止の必要なこともこれに劣らない。蚕種関係者がウイルスの経卵伝達に関心を特つのは、蚕種の内部にかくれた汚染の警戒のためであるが、内部汚染の起こるような状況の下では、必らず卵の外部の汚染も伴なう筈であり、更に外部汚染は感染し得るウイルスであるから、これが次代蚕におよぼす影響は上に述べた程度の内部汚染以上に恐ろしく、見かけの上では経卵伝達と同じ結果になる。
伝染性軟化病ウイルスは、鮎沢ら(19)によれば比重1.075の塩酸液ならば常温5分間の浸漬によってほぼ完全に不活化され、渡辺ら(1152)によれば比重1.110の塩酸液では24℃、1時間、または48℃、6分間で完全に不活化されると云う(渡辺らは比重1.110の塩酸液を濃度15%と記している。比重と濃度との何れが誤植かは不明である)。同じウイルスは、鮎沢らによれは400倍、10分間、渡辺らによれば200倍、10℃、15分間のクライト液浸漬によって完全に不活化する。
内海ら(1129)はβ-プロピオラクトンが卵面消毒にも有効なことを報告しているが、実用上の問題としてはなお検討すべき点があるように思われる。
多角体に封入されているウイルスは伝染性軟化病ウイルスよりは抵抗力が強いが、ホルマリンガスによって卵面消毒を完全に行ない得ることが知られている(Ⅸ1C)。
しかし、最も大切なことは、このような単なる卵面消毒の技術ではなく、何処に汚染の原因があるかに常に注意して、飼育から蚕種の調製までの総てを通じ、清浄な蚕種製造に心掛けることである。いくら卵面だけをきれいにしても容器、保護室、冷蔵庫、催青室、採種室、蚕種庫などが汚れておれば意味はない。
例えば、採種室、冷蔵庫、人工孵化後の乾燥室などから、埃を集めて滅菌蒸留水で浸出し、その濾液を桑に塗って蟻蚕に与えた処、伝染性軟化病が発生したとか(1181)、蚕種製造室の雄部屋の鱗毛を掃立桑にまぶして1回(15時間)添食した蚕に伝染性軟化病が多発した(310)などの報告はこの危険を物語っているもので、器具や作業員を通して卵の汚染する機会は少なくない。
また、外観上全く健康な蚕(1180)や正常に産卵した蛾(736,1033)が感染性のウイルスを保有していることも多いから、飼有中に病蚕が出なかった場合にも、感染や汚染のおそれがある。
対策の第二は卵、幼虫、蛹、蛾の各期を通じて取扱を適正にすることである。蚕が潜在性ウイルスを持っていても、これを活性化する誘発条件が働かなければ発病することはないのであり、幸なことに、ウイルス病の誘発条件になると云われている不良環境は、蚕種の保護取扱上悪いことの知られているものばかりであるから、蚕種の保護、取扱いを適正に行なうことが直ちに対策に連ることになる。
直接にウイルス病を誘発する条件にはならなくても、不良環境の下で採種した卵から孵化した蟻蚕がウイルス病に感染し易い(Ⅱ2Ac)ことにも注意する必要がある。
著者名はヘボン式綴りが多いように思われるので、大体その順序で配列した。しかし、HASIMOTO、HUKUHARA、ITIKAWA、KOIDSUMIなど、わかっている限りはそれぞれの綴りに従った。また、漢字の読み方のわからない著者名もあるので、配列は必らずしも統一されていない。*を付けた論文は原著をみていないものである。外国の文献は挙げない。
1)阿部文作(1952) 日蚕雑 21(2・3),123
2)阿部泰治(1949) 日蚕東北 (2),51-52
3)阿部泰治,高橋正(1950) 日蚕東北 (3),89-90
4)阿部泰治,八鍬春美,篠木順二,山科ヨシミ(1964) 日蚕雑 33(1),97-98
5)阿部泰治,柳谷恒夫(1961) 日蚕東北 (15),2
6)足立豪男,渡辺昭典,近繩清文,竹野逸夫(1959) 日蚕九州(昭34),18
7)足立豪男,渡辺昭典,新倉克己(1959) 日蚕九州(昭34),18
8)足立豪男,渡辺昭典,新倉克己(1961) 日蚕九州(昭35),2-3
9)足立豪男,渡辺昭典,竹野逸夫(1959) 日蚕九州(昭34),19
10)足立豪男,渡辺昭典,竹野逸夫(1961) 日蚕九州(昭35),1-2
11)足立豪男,渡辺昭典,竹野逸夫(1961) 日蚕九州(昭35),2
12)足立豪男,渡辺昭典,竹野逸夫,近繩清文(1961) 日蚕九州(昭35),3
13)足立豪男,渡辺昭典,竹下タヨ(1961) 日蚕九州(昭35),1
14)足立雅夫(1953) 柏原製糸調査資料(6),1-5
15)足立美佐男(1926) 群馬蚕試報 (1),1-24
16)足立美佐男(1947) 日蚕東北 (1),34-36
17)愛知県蚕業試験場(1954) 愛知蚕試彙 (13),1-73
18)鮎沢啓夫,古田要二,倉田啓而(1963) 日蚕雑 32(3),185
19)鮎沢啓夫,古田要二,倉田啓而,佐藤文子(1964) 蚕試報 19(2),223-240
19a)鮎沢啓夫,古田要二,中村晃三(1961) 日蚕雑 30(5),405-412
20)鮎沢啓夫,倉田啓而(1964) 日蚕雑 33(3),242,243
21)鮎沢千尋(1961) 日蚕雑 30(3),238
23)鮎沢千尋,古田要二,鮎沢啓夫(1962) 日蚕雑 31(3),194
24)安喰定洽(1950) 日蚕東北 (3),97-99
25)味岡秀夫,十万五良(1937) 鹿児島蚕試報 (23),136-142
26)赤羽武夫(1940) 蚕業新報 48,566
27)赤井 弘(1956) 日蚕雑 25(3),214
28)赤井 弘(1957) 日蚕雑 26(5),335-340
29)赤井 弘(1958) 蚕糸研究 (26),19-21
30)赤井 弘(1958) 日蚕雑 27(6),403-408
31)赤尾 晃(1935) 動雑 47(557),159-163
32)赤尾 晃(1941) 蚕試報 10(3),189-227
33)赤尾 晃(1942) 日蚕雑 13(3),110-113
34)赤尾 晃(1943) 蚕試報 11(3),295-309
35)赤尾 晃(1943) 日蚕雑 14(1・2),113-118
36)赤尾 晃(1944) 蚕試報 12(1),1-16
37)AKAO,A.(1939) Jour.Biochem. 30(2),303-349
38)明石弘(1908) 日本昆虫学会々報 2(6),145-146
39)秋田儀三郎(1947) 日蚕東北 (1),54-57
40)秋田康一,小林春雑(1955) 東京都立大理研究要録 (5),36
41)雨宮 平(1957) 蚕糸研究 (20),19-21
42)雨宮 平,提 要造(1953) 蚕糸研究 (4),5-10
43)雨宮 平,提 要造(1954) 蚕糸研究 (10),26-27
44)雨宮 平,提 要造,藤巻忠彦(1956) 蚕糸研究 (16),31-39
45)雨宮 平,提 要造,藤巻忠彦(1957) 蚕糸研究 (19),5-8
46)AOKI,K. (1962) Sci.Rept.TohokuUniv.Ⅳ Ser.(Biol.) 28(1),29-36
47)荒井成彦,伊藤智夫(1963) 蚕試報 18(4),231-250
48)荒木武雄,三田伊三郎,三浦英太郎(1912) 京都蚕講成績 (25),117-131
49)荒木武雄,三田伊三郎,三浦英太郎(1912) 京都蚕講成績 (25),221-235
50)荒木武雄,三田伊三郎,三浦英太郎(1913) 京都蚕講成績 (26),1-50
51)荒木武雄,三田伊三郎,渡辺準太郎,三浦英太郎(1910) 京都蚕講報 (19),17-144
52)荒木武雄,三浦英太郎(1912) 京都蚕講成績 (25),133-135
53)荒木武雄,三浦英太郎(1912) 京都蚕講成績 (25),237-242
54)荒木武雄,三浦英太郎(1912) 京都蚕講成績 (25),243-246
55)荒木武雄,三浦英太郎(1913) 京都蚕講成績 (26),217-240
56)荒木武雄,三浦英太郎(1913) 京都蚕講成績 (26),241-257
57)荒木武雄,三浦英太郎(1913) 京都蚕講成績 (26),259-341
58)荒木武雄,三浦英太郎(1914-16) 大日本蚕糸会報 (274),1-17
; (275),28-31 ;
(276),21-26 ; (277)*;(281),598-603;(282),689-693
; (283),774-776 ;
(284),879-881;(285),971-972;(286),1075-1077
; (287),1159-1161 ;
(288),49-53;(289),153-156;(290),238-242
59)荒木弥蔵(1926) 衣笠蚕報 (233),44-49
60)有賀久雄(1938) 日蚕雑 9(3),275-276
61)有賀久雄(1957) 日蚕雑 26(2),9-15
62)有賀久雄,福原敏彦(1961) 日蚕雑 30(4),334-338
63)有賀久雄,金井栄一,アムヌイ・イサランクール・ナ・アユタヤ(1959) 日蚕雑 28(6),369-374
64)有賀久雄,長島栄一,武井隆三(1964) 日蚕雑 33(6),460-463
65)有賀久雄,吉武成美(1957) 日蚕雑 26(1),21-30
66)有賀久雄,吉武成美,江口正治(1952) 日蚕雑 21(5・6),255-263
67)ARUGA,H. and NAGASHIMA,E.(1962) Jour.Ins.Pathol.4(3),313-320
68)芦田良吉(1928) 衣笠蚕報 (264),244-248
69)芦田良吉,浜ノ上久福(1937) 和歌山蚕試彙 (1),21-30
70)ASTAUROV,B.L.(1967) 日蚕雑 36(4),277-285
71)尾藤省三(1931) 応動 3(5),314-318
72)地方蚕業試験場協力試験(年号なし) 技術資料 (43),1-3
73)地方蚕業試験場協力試験(1960) 技術資料 (48),1-7
74)地方蚕業試験場協力試験(1960)技術資料 (48),11-17
75)筑紫春生(1952) 田中義麿:家蚕遺伝学 第Ⅲ章.裳華房,東京
76)茅野春雄(1958) 科学 28(5),233-238
77)茅野春雄(1959) 実験形態学誌 (13),13-19
78)茅野春雄(1960) 日蚕雑 29(3),270
79)茅野春雄(1962) 生化学 34(6),235-242
80)CHlNO,H.(1956) Embryologia 3(2),167-186
81)CHlNO,H.(1957) Embryologia 3(4),295-316
82)CHlNO,H.(1958) Jour.Ins.Physiol. 2,1-12
83)CHlNO,H.(1960) Jour.Ins.Physiol. 5,1-15
84)CHlNO,H.(1961) Jour.Ins.Physiol. 6,231-240
85)CHlNO,H.(1963) Arch.Biochem.Biophysics 102,
400-415
86)小県金策(1932) 衣笠蚕報(308),13-19
87)土井良宏(1968) 九大農学芸 23(4),205-214
88)道家信道(1935) 応動 7(2),63-69
89)江ロ正治(1955) 日蚕雑 24(5・6),344-349
90)江ロ正治(1955) 日蚕雑 24(5・6),350-352
91)江ロ正治,政山 亨,西村允子(1966) 日蚕雑 35(6),435-443
92)江ロ嘉清(1954) 蚕糸研究 (8),10-13
93)江ロ嘉清,穂之上但,和田武春(1963) 蚕糸研究(47),26-29
94)江ロ嘉清,中田昌保,飛田 勉(1954) 蚕糸研究 (8),3-9
95)江竜寛一(1942) 日蚕雑 13(3),148-149
96)江竜寛一(1943) 日蚕雑 14(5・6),311-319
97)藤本直正(1943) 日蚕雑 14(5・6),283-293
98)藤本直正(1951) 日蚕雑 20(4),258-263
99)藤本直正(1956) 日蚕雑 25(2),103-110
100)藤本直正(1958) 日蚕雑27(3),111-116
101)藤本直正(1958) 実験形態学誌 (12),94
102)藤原 公(1954) 蚕糸研究 (10),22-25
103)藤原 公(1955) 蚕糸研究 (12),5-8
104)藤原 公,大柳正之(1956) 蚕糸研究 (15),39-41
105)福田仁郎(1936) 応動8(3),116-123
106)福田正人(1936) 衣笠蚕報 (360),440-442
107)福田宗一(1942) 動雑 54(4),156-157
108)福田宗一(1948) 日蚕雑 17(3・4),113
109)福田宗一(1949) 日蚕雑 18(5),313
110)福田宗一(1949) 日蚕雑 18(5),314
111)福田宗一(1951) 動雑 60(6),119-121
112)福田宗一(1953) 日蚕中部 (6),1-15
113)福田宗一(1954) 動雑 63(11・12),468
114)福田宗一(1955) 実験形態学誌 (9),22-27
115)福田宗一(1956) 科学 26(3),118-124
116)福田宗一(1960) 日蚕雑 29(3),287
117)福田宗一,日高敏隆(1959) 実験形態学新説(竹脇 潔・針塚正樹・深谷昌次編)第1編Ⅰ,養賢堂、東京
118)福田宗一,櫛田満一(1949) 日蚕雑 18(2),88-94
119)福田宗一,河野太郎(1952) 日蚕雑 21(2・3),124-125
120)福田宗一,河野太郎(1953) 日蚕雑 22(3),122
121)福田宗一,河野太郎(1954) 日蚕雑 23(3),181
122)福田宗一,河野太郎(1955) 日蚕雑 24(3),188
123)福田宗一,河野太郎(1955) 日蚕雑 24(3),188
124)福田宗一,河野太郎(1956) 日蚕雑 25(3),221
125)福田宗一,河野太郎(1956) 日蚕雑 25(6),389-394
126)福田宗一,河野太郎(1957) 日蚕雑 26(3),219
127)福田宗一,河野太郎(1958) 日蚕雑 27(3),156
128)福田宗一,河野太郎(1959) 日蚕雑 28(3),164
129)福田宗一,河野太郎(1960) 日蚕臨時大会論演集(中部),11
130)福田宗一,酒井 績(1956) 日蚕雑 25(3),221-222
131)福田宗一,酒井 績(1957) 動雑 66(2・3),66-67
132)FUKUDA,S.(1951) Proc.Japan Acad.27(10),672-677
133)FUKUDA,S.(1952) Annoy.Zool.Japon. 25(1・2),149-155
134)FUKUDA,S.(1953) Proc.Japan Acad.29(7),381-384
135)FUKUDA,S.(1953) Proc.Japan Acad.29(7),385-388
136)FUKUDA,S.(1953) Proc.Japan Acad.29(7),389-391
137)FUKUDA,S.and KOHNO,T.(1964) Embryologia 8(3),177-190
138)FUKUDA,S.and TAKEUCHI,S.(1967) Proc.Japan Acad.43(1),51-56
139)FUKUDA,S.and TAKEUCHI,S.(1967) Embryologia 9(4),333-353
139a)福田紀文(1960) 蚕試報 15(10),595-610
140)福田紀文,亀山多美子,松田基一(1963) 蚕試報 18(3),157-171
141)福島県蚕業試験場(1927) 福馬蚕試報 (1),156-197
142)古田要二(1968) 蚕糸研究 (68),128-132
143)二木猪一(1952) 日蚕東北 (5),11-13
144)二木猪一(1952) 技術資料 (33),1
145)蒲生卓磨(1960) 信大繊維報 (10),124-129
146)蒲生卓磨(1965) 蚕試報 19(3),255-307
147)蒲生俊興,竹田 寛,塩入 勤(1949) 技術資料(13),33
148)浜 次雄(1936) 応動8(6), 318-327
149)浜崎信太郎(1930) 応動 2(3),165-170
150)原田忠次,真野保久(1952) 蚕糸研究 (2),29-32
151)針塚正樹(1966) 日蚕雑 35(4),307-308
152)長谷川金作(1938) 日蚕雑 9(3),282-283
153)長谷川金作(1943) 日蚕雑14(1・2),24-32
154)長谷川金作(1943) 蚕試報 11(3),359-377
155)長谷川金作(1947) 蚕試報 12(5),481-492
156)長谷川金作(1947) 蚕試報 12(5),493-512
157)長谷川金作(1947) 蚕試報 12(5),513-530
158)長谷川金作(1951) 日蚕雑 20(6),403-406
159)長谷川金作(1952) 動雑 61(3・4),82
160)長谷川金作(1952) 日蚕雑 21(1),1-6
161)長谷川金作(1952) 日蚕雑 21(2・3),95-96
162)長谷川金作(1955) 日蚕東海(3),25-30
163)長谷川金作(1957) 日蚕雑 26(3),232
164)長谷川金作(1959) 実験形態学新説(竹脇 潔・針塚正樹・深谷昌次編)第1編Ⅲ.養賢堂,東京
165)長谷川金作(1963) 日蚕雑 32(6),367
166)HASEGAWA,K.(1951) Proc.Japan Acad. 27(10),667-671
167)HASEGAWA,K.(1952) Jour.Fac.Agr.Tottori Univ. 1(2),83-124
168)HASEGAWA,K.(1957) Nature 179, 1300-1301
169)HASEGAWA,K.(1963) J.Exp.Biol. 41,517-529
170)HASEGAWA,K.(1964) J.Exp.Biol. 41,855-863
171)HASEGAWA,K. and YAMASHITA,O.(1965) J.Exp.Biol. 43,271-277
172)HASEGAWA,K. and YAMASHITA,O.(1967) 日蚕雑36(4),297-301
173)橋田久三郎,荒尾照夫(1962) 日蚕関西 (24),7
174)橋爪貞三(1955) 蚕糸研究 (14),21-23
175)橋本春雄(1930) 日蚕雑1(2),151-153
176)橋本春雄(1934) 蚕試報 8(10),455-464
177)橋本春雄(1948) 日蚕雑 16(3・4),62-64
178)橋本春雄(1951) 日蚕雑 20(2),164-165
179)橋本春雄(1953) 日蚕雑 22(4),175-180
180)橋本春雄(1953) 日蚕雑 22(5),205-210
181)橋本春雄(1956) 日蚕雑 25(3),215-216
182)橋本春雄(1956) 遺雑 31(10 ・ 11),294
183)橋本春雄(1957) 蚕糸研究 (20),11-13
184)橋本春雄(1961) 日蚕雑 30(2),78-82
185)橋本春雄,百瀬 絢,久保村安衛(1959) 日蚕雑28(4),199-203
186で)秦 信親,広部達道外11名(1964) 日蚕雑 33(3),259-260
187)秦 専章(1924) 蚕業新報 32(367),15-26
188)服部文雄(1929) 群馬蚕試報 (7),1-152
189)服部達治,四方正義(1961) 日蚕雑 30(3),246
190)林禎二郎(1937) 九大農学芸 7(4),359-372
191)樋口太郎(1920) 佐久良会誌 (7),40-45
192)引地栄一(1960) 日蚕東北 (14),27-28
193)引地栄一(1961) 日蚕東北 (15),15
194)引地栄一(1962) 岩手蚕試年報 (9),93-95
195)平野茂市(1953) 動雑 62(3・4),76
196)平野茂市(1954) 動雑 63(3・4),106
197)平野茂市(1960) 動雑 69(9),292-295
198)平野茂市(1961) 動雑 70(9),316-322
199)平尾常男,五十嵐三郎(1956) 蚕糸研究 (16),12-15
200)平山正生(1950) 日蚕東北 (3),96-97
201)平山正生,黒沢六良(1951) 日蚕東北 (4),53-55
202)平山正生,館石正助(1952) 日蚕東北 (5),21-23
203)広部達道(1960) 日蚕雑 29(3),277-278
204)広部達道,中島文人(1967) 日蚕雑 36(3),253-254
205)広部達道,岡田一次,下口曠恩(1964) 日蚕雑 33(3),231
206)広部達道,大井秀夫,皆川 勇(1953) 蚕糸研究 (4),14-16
207)広部達道,大井秀夫,皆川 勇(1956) 日蚕雑 25(3),218-219
208)広部達道,大井秀夫,皆川 勇,生方せつ(1956) 日蚕雑 25(6),375-381
209)広部達道,大井秀夫,皆川 勇,生方せつ(1956) 日蚕雑 25(6),382-388
210)広部達道,矢崎忠義(1966) 日蚕雑 35(3),223
211)広部達道,矢崎忠義(1966) 日蚕雑 35(3),223
212)菱田昭夫,林 孝之(1962) 日蚕雑 31(2),69-72
213)本間正司(1958) 日蚕関東 (10),17
214)本多岩次郎(1904) 大日本蚕糸会報 (140),4-6
215)堀 国彦(1956) 愛知蚕試概要(昭30),67-71
216)堀口宜之(1930) 蚕業新報 38(440),204-210
217)堀内彬明,波島千恵子,安江 昇(1962) 日蚕雑 31(6),421-423
218)堀内彬明,昆野民夫,波島千恵子(1959) 蚕糸研究 (30),4-7
219)堀内彬明,中村晃三,波島千恵子(1960) 蚕糸研究 (34),9-13
220)堀内彬明,入戸野康彦,功力文良,長池幸男(1965) 蚕試彙 (85),1-25
221)堀内彬明,清水 滋(1960) 蚕糸研究 (35),18-20
222)堀内彬明,清水 滋(1965) 蚕糸研究 (55),27-32
223)堀内彬明,清水 滋,波島千恵子,安江 昇(1964) 日蚕雑 33(1),63-66
224)堀内彬明,清水 滋,中村晃三,安江昇(1961) 蚕糸研究 (37),24-27
225)堀内彬明,清水 滋,安江 昇(1964) 日蚕雑 33(1),94-95
226)堀内彬明,清水 滋,安江 昇(1965) 日蚕雑 34(3),177-178
226a)星沢康鎮(1930) 蚕糸界報 39(461),864-219
228)堀田成一郎,河端常信(1955) 岩手蚕試年報 (2),2215
229)福原敏彦(1962) 日蚕雑 31(2),97-100
230)兵藤義幸(1924) 大日本蚕糸会報 33(393),817-823
231)兵藤義幸(1924) 佐久良会誌 (16),7-15
232)兵藤義幸(1925) 大日本蚕糸会報 34(396),134-138
; (397),221-225
233)兵藤義幸,松井護郎(1923) 愛媛蚕試報 (11),32-37
234)維田 潔(1943) 蚕糸界報 52(616),7-13
235)五十嵐三郎(1960) 蚕糸研究 (35),1-5
236)五十嵐三郎,杉山八郎(1964) 蚕糸研究 (49),42-52
237)池田栄太郎(1910) 大日本蚕糸会報 (221),6-8 ; (222),7-9
; (223),10-12 ; (224),8-10
238)池田栄太郎(1913) 実験蚕体解剖生理論.明文堂,東京
239)池田栄太郎(1915) 大日本蚕糸会報 24(276),13-16
240)池田次男(1947) 日蚕東北 (1),63-65
241)池田次男(1949) 日蚕雑 18(3),161-168
242)池江輝二(1924) 長崎蚕試報(6),1-94
243)稲神 馨(1954) 日蚕雑 23(4),228-232
244)稲神 馨(1955) 日蚕雑 24(2),91-96
245)稲神 馨(1955) 日蚕雑 24(3),195
246)稲神 馨,須藤芳三(1955) 日蚕雑 24(2),85-91
247)入来重盛(1941) 動雑 53(2),123-124
248)入来重盛,平野茂市(1952) 動雑 61(3・4),98-99
249)石井好一(1960) 日蚕雑 29(3),256
250)石井弥平(1933) 日蚕雑 4(2),74-82
251)石川金太郎(1926) 動雑 38(450・451),104-114
252)石川金太郎(1928) 蚕糸界報 37(432),210-217
253)石川金太郎(1933) 蚕糸界報 42(497),45-46
254)石川金太郎(1933) 応動 5(5),243-244
255)石川金太郎(1952) 技術資料 (33),2-3
256)石川美之利,佐野利男,鎌倉今朝五郎(1966) 静岡蚕試報 (17),53-57
257)石川美之利,佐野利男,鎌倉今朝五郎(1967) 静岡蚕試報 (18),22-26
258)石川 統(1965) 動雑 74(11・12),336
258a)石森直人(1942) 蚕.岩波書店,東京
259)石渡繁胤(1904) 蚕業新報 (134),456-457
260)石渡繁胤(1913) 大日本蚕糸会報 (256),11-12 ;(257)*;(258),14-15
; (259),10-14
261)石渡繁胤(1913) 蚕業新報 21(244),21-23;(245),34-38
262)石渡繁胤(1926) 衣笠蚕報 8(6),421-433
263)ISHIWATA,S.(1904) 大日本蚕糸会報 (146),1-2
260)磯谷 鋭(1941) 蚕種製造実務要覧.明文堂,東京
265)板谷健吾(1936) 蚕体生理学.明文堂,東京
266)板谷健吾(1940) 日蚕雑 11(3),113-140
267)市川信一(1951) 日蚕報 20(4),297
268)市川信一(1952) 蚕試報 14(2),23-91
269)市川信一(1952) 日蚕報 21(5・6),289
270)市川信一(1955) 日蚕報 24(3),214
271)市川信一(1956) 日蚕雑 25(3),214-215
272)市川信一,剣持謙二(1959) 蚕糸研究(32)1-6
273)伊藤盛次,黒沢益雄(1931) 衣笠蚕報 (300),37-46
274)伊藤盛次,黒沢益雄(1932) 衣笠蚕報 (307),1-12
275)伊藤盛次,黒沢益雄(1932) 衣笠蚕報 (312),21-43
276)伊藤盛次,黒沢益雄(1933) 衣笠蚕報 (318),1-16
277)伊藤智夫(1950) 日蚕雑 19(6),523-525
278)伊藤智夫(1950) 日蚕雑 19(6),526-529
279)伊藤智夫(1952) 蚕糸研究 (2),157-158
280)伊藤智夫(1954) 蚕試報 14(5),263-278
281)伊藤智夫,荒井政彦(1963) 蚕試報 18(4),209-229
282)伊藤智夫,堀江保宏,渡辺喜二郎 外2名(1966) 日蚕雑 35(5),374-379
283)伊藤智夫,田中元三(1952) 蚕糸研究 (2),25-28
284)伊藤智夫,田中元三(1953) 蚕糸研究 (3),26-29
285)伊藤豊雄(1962) 蚕糸研究 (43),38-42
286)伊藤豊雄,松野道雄,渡辺一己(1965) 蚕糸研究 (55),11-18
287)岩野関寿郎,西尾秋雄,赤沢日出子(1963) 徳島蚕試蚕糸技術 10(1),6-11
288)岩崎行高(1929) 蚕糸界報 (453),1378
289)岩崎行高(1931) 応動 3(5),308-313
290)岩崎行高(1932) 鹿児島高農報 (10),191-195
291)岩崎行高(1932) 鹿児島高農報 (10),197-201
292)岩崎行高,小野二郎(1929) 蚕糸界報 (453),1378-1381
293)岩崎行高,小野二郎(1929) 蚕業新報 37(437),1294-1296
294)岩下嘉光(1955) 日蚕雑 24(3),198-199
295)伊与田茂,米山好人(1933) 日蚕雑 4(3),193-209
290 地引嘉作,高橋一太(1952) 日蚕中部 (4),2-3
297)十万五良(1961) 日蚕関西 (23),28-29
298)十万五良(1963) 日蚕関西 (26),20
299)十万五良,片岡二郎(1954) 高知蚕試蚕桑彙 (2),17-18
300)十万五良,岸本稔(1958) 高知蚕試蚕桑彙 (6),24
301)樺沢 敦,瀬戸川喜多夫,蛯原富男(1960) 日蚕雑 29(3),261
302)鎌倉今朝五郎(1962) 日蚕雑 31(3),188
303)鎌倉今朝五郎(1966) 静岡蚕試報 (17),47-53
304)鎌倉今朝五郎,金刺みか江(196) 1静岡蚕試報 (12),30-38
305)鎌倉今朝五郎,鈴木実(1960) 日蚕東海 (8),22-23
306)上条太八郎,井出 智(1965) 日蚕雑 34(3),200-201
307)金井 保(1959) 日蚕関西 (19),21
308)加納繁隆,原 孫一,小栗敏彦(1962) 岐阜蚕試彙 (5),81-85
309)笠井隆善(1932) 蚕糸界報 41(490),55-63
310)片岡 平,梅村義雄(1965) 日蚕雑 34(3),213
311)加藤 久(1960) 蚕糸研究 (34),4-8
312)加藤 久,叶内朝治,土屋久一(1952) 日蚕東北 (5),13-14
313)加藤 久,笹原重雄,佐藤喜美雄(1967) 蚕糸研究 (62),19-21
314)加藤 久,富樫きん(1961) 日蚕東北 (15),2-3
315)加藤 勝(1952) 日蚕雑 21(5・6),252
316)加藤和一郎(1915) 大日本蚕糸会報 24(282),693-696
317)勝又藤夫(1928) 蚕糸雑 1(1),1-11
318)勝又藤夫(1954) 柏原製糸調査資料 (9),1-9
319)KATSUMATA,F.(1968) 日蚕雑 37(6),453461
320)勝野貞哉(1965) 日蚕雑 34(3),203
321)勝野貞哉(1966) 日蚕雑 35(3),227
322)勝野貞哉(1967) 日蚕雑 36(3),249
323)川畑外志夫(1960) 日蚕雑 29(3),266-267
324)河端常信(1956) 岩手蚕試年報 (3),16-17
325)川口栄作(1926) 蚕業新報 34(391),38-41
326)川口栄作(1926) 動雑 38(447),11-13
327)川口栄作(1933) 日蚕雑 4,172-173
328)川口栄作(1933) 日蚕雑 4,173-175
329)川口栄作(1934) 日蚕雑 5(1),1-20
330)川口栄作(1938) 遺雑 14(6),262-263
331)川口栄作,宮慶一郎(1943) 遺雑 19(3),133-134
332)川口栄作,滝沢義郎(1942) 日蚕雑 13(3),133-135
333)川口栄作,滝沢義郎(1942) 日蚕雑 13(3),135-138
334)KAWAGUCHI,E.(1928) Zeit.Zell.mikr.Anat. 7(4),519-552
335)河合金次郎,江連恒夫(1953) 千葉蚕試報 (3),1-17
336)河合 孝(1950) 日蚕雑 19(3),207-212
337)河合 孝(1951) 日蚕雑 20(4),288
338)河合 孝(1951) 日蚕雑 20(6),413-417
339)河合 孝(1952) 日蚕雑 21(5・6),253-254
340)河合 孝(1954) 日蚕雑 23(3),147-150
341)河合 孝(1956) 日蚕雑 25(2),111-114
342)河合 孝(1956) 日蚕雑 25(3),222-223
343)河合 孝(1956) 日蚕雑 25(4),263-267
344)河合 孝(1957) 日蚕雑 26(2),55-59
345)河合 孝(1957) 日蚕雑 26(2),60-64
346)河合 孝(1958) 日蚕雑 27(1),30-33
347)河合 孝(1958) 日蚕雑 27(3),117-121
348)河合 孝(1958) 日蚕雑 27(3),156
349)河合 孝(1959) 日蚕雑 28(6),358-361
350)河合 孝(1960) 日蚕雑 29(4),345-348
351)河合 孝(1960) 日蚕雑 29(4),349-351
352)河合 孝(1961) 日蚕雑 30(3),197-200
353)河合 孝(1961) 日蚕関西 (23),28
354)河合 孝(1962) 応動昆 6(1),24-26
355)河合 孝(1962) 日蚕雑 31(3),190
356)河合 孝(1962) 鳥取農学会報 14,8-15
357)河合 孝(1962) 鳥取農学会報 14,16-21
358)河合 孝(1963) 日蚕雑 32(3),192
359)河合 孝,江花哲也(1954) 鳥取農学会報 10(3),95-98
360)河北金太郎(1947) 日蚕東北 (1),40-42
361)河北金太郎、河田明二(1956) 日蚕関西 (13),21
362)河倉義安(1927) 漁農科学 1(1),14-26 ; (5),1-13
363)川浪 中(1950) 日蚕雑 19(3),255-269
364)川瀬茂実(1954) 日蚕雑 23(5),296-298
365)川瀬茂実(1956) 日蚕雑 25(4),257-262
366)川瀬茂実(1956) 日蚕雑 25(5),322-326
367)川瀬惣次郎,岩岡末彦,斎藤良三郎(1913) 農学会報 (136),21-41
368)川瀬惣次郎,松林竹比虎(1917-18) 大日本蚕糸会報 (311),986-996;(312),44-48;(313),135-140
369)吉川秀男(1940) 日蚕雑 11(4),227-234
370)吉川秀男(1942) 動雑 54(7),245-249
371)吉川秀男(1943) 蚕試報 11(3),311-345
372)吉川秀男(1951) 日蚕報 20(4),292-293
373)KIKKAWA,H.(1953) Advances in Genetics 5,107-140
374)菊地六郎(1951) 技術資料 (29),67
375)菊地六郎(1953) 日蚕雑 22(3),122-123
376)金原和一,北爪雄次郎(1956) 群蚕要報 (30),28-35
377)木村真作(1952) 日蚕雑 21(2・3),149-150
378)木村真作(1953) 日蚕雑 22(3),131-132
379)木村真作,水出通男(1952) 蚕糸研究 (71),69-74
380)木下 実(1951) 蚕糸界報 60,(698)13-18
381)木下 実(1951) 技術資料 (29),21
382)木下 実(1952) 技術資料 (33),33
383)木下 実(1956) 神奈川県蚕試要報 (2),60-62
383a)岸田 豊(1931) 蚕糸界報 40(473),877-879
384)北沢周一(1937) 蚕糸界報 46(546),42-48
385)北沢敏男(1957) 日蚕雑 26(73),231-232
386)北沢敏男,神田俊男,高見丈夫(1963) 蚕試報 8(5),283-295
387)北沢敏男,高見丈夫(1963) 蚕試彙 (81),1-4
387a)北沢敏男,高見丈夫(1967) 日蚕関東 (18),36
388)小原隆三(1967) 応動昆 11(2),71-75
389)小針喜三郎(1924) 実験蚕種の人工孵化法.蚕糸雑誌社,上田
390)小針喜三郎,金子哲郎(1938) 日蚕雑 9(2),125-143
391)小針喜三郎,金子哲郎(1939) 日蚕雑 10(3),212-213
392)小針喜三郎,室賀兵左衛門(1926) 蚕業新報 34(401),1252-1257
; (402),1379-1384
393)小針喜三郎,室賀兵左衛門(1927) 蚕業新報 35(408),686-691
394)小針喜三郎,室賀兵左衛門(1927) 蚕業新報 35(411),1069-1078
395)小針喜三郎,室賀兵左衛門(1930) 蚕業新報 38(447),1180-1185
396)小針喜三郎,室賀兵左衛門(1932) 蚕業新報 (462),22-27
397)*小針喜三郎,室賀兵左衛門(1932) 片倉蚕業試験所報告 1(1)
398)小針喜三郎,室賀兵左衛門,丸山徳雄(1929) 蚕業新報 37(431),591-598
399)KOBAYASHI,H. and AKITA,Y.(1962) Embryologia 7 (1),1-11
400)小林勝利(1956) 日蚕雑 25(3),232
401)小林勝利(1957) 蚕試報 15 (3),181-273
402)小林勝利,石戸谷幸雄,楠木園正雄(1966) 日蚕雑 35(3),217-218
403)小林勝利,楠本薗正雄(1965) 日蚕雑 34(3),218
404)小林勝利,横平喜代子(1964) 日蚕雑 33(3),255-256
405)KOBAYASHI,T.(1962) Ann.Rept.Nat.Inst.Gen.Japan (12),91-92
406)木暮槇太(1930) 長野蚕試報 (11),1-152
407)木暮槇太(1935) 原蚕飼育の要諦.大日本蚕糸会,東京
408)木暮槇太,山本茂樹(1930) 日蚕雑 1(1),23-47
409)KOGURE,M.(1933) Jour.Dept.Agr.Kyushu Imp. 4(1),1-93
410)小泉清明,和田吉弘(1955) 応動 20(4),184-190
411)小池弘三(1914) 大日本蚕糸会報 (273) 24-27 ;
(274),34-37
412)小池利男,飛田永男(1954) 長野蚕試蚕糸技術 17(10),219-238
413)小池勇次郎,土屋久一(1951) 日蚕雑 20(1),73
414)小池勇次郎,土屋久一,森 文蔵(1949) 日蚕東北 (2),40-42
415)小池勇次郎,土屋久一,森 文蔵(1949) 日蚕東北 (2),76-78
416)小池勇次郎,土屋久一,森 文蔵(1951) 日蚕東北 (4),87-88
417)小岩 □,小針要吉(1965) 蚕糸研究 (55),6-10
418)小岩井宗作(1910) 大日本蚕糸会報 (215),10-14;(217),18-21
; (218),16-18
419)小泉二郎(1957) 蚕糸研究 (23),1-11
420)小泉二郎(1958) 蚕糸研究 (24),1-6
421)小泉二郎(1958) 蚕糸研究 (25),3649
422)-1958蚕糸研究(27),16-23
423)小泉二郎,橋詰 強,小針洋子(1963) 蚕糸研究 (45),9-11
424)小泉二郎,橋詰 強,小針洋子(1964) 蚕糸研究 (49),31-34
425)小泉二郎,橋詰 強,高野幸治,小針洋子(1964) 蚕糸研究 (49),35-38
426)小泉二郎,小針洋子(196) 蚕糸研究 (37),11-14
427)小泉二郎,久保田重良,松田洋子(1959) 蚕糸研究 (32),22-23
428)小泉二郎,久保田重良,松田洋子(1960) 蚕糸研究 (33),19-21
429)小泉二郎,久保田重良,松田洋子(1961) 蚕糸研究 (37),29-36
430)小泉二郎,松田洋子,入戸野康彦外3名(1960) 蚕試彙 (77),41-54
431)小泉二郎,塩見昭男,小針洋子(1961) 蚕糸研究 (39),1-4
432)小泉二郎,塩見昭男,小針洋子(1962) 蚕糸研究 (40),7-10
433)小泉二郎,塩見昭男,小針洋子(1962) 蚕糸研究 (40),11-18
434)小泉二郎,塩見昭男,小針洋子(1962) 蚕糸研究 (40),19-20
435)小泉二郎,高野幸治,柳川弘明(1965) 蚕糸研究 (55),19-21
436)古和田敬治(1950) 技術資料 (22),10-11
437)古和田敬治(1960) 日蚕関西 (22),7
438)古和田敬治(1961) 日蚕雑 30(3),247
439)古和田敬治(1962) 日蚕雑 31(3),191
440)古和田敬治(1963) 日蚕関西 (26),21
441)古和田敬治(1963) 日蚕関西 (27),2
442)古和田敬治,荒木嘉市,菅野 勝(1962) 日蚕関西 (24),9-10
443)小山長雄,滝沢達夫(196) 日蚕雑 31(3),192
444)KOYAMA,N. and FURIHATA,Y.(1965) 日本生態学会誌 15(3),89-93
445)高知県原蚕種製造所(1917) 高知県原蚕事績報 (1),58-60
446)河野茂盛(1925) 蚕業新報 33(383),516-525
; (384),646-654
447)河野太郎(1951) 技術資料 (29),19
448)河野太郎(1960) 日蚕雑 29(3),314-315
449)河野太郎(1961) 日蚕雑 30(3),249
450)河野太郎(1962) 片倉工業蚕業研報 (特別2),1-81
451)河野太郎(1962) 日蚕雑 31(3),189
452)河野太郎(1964) 日蚕雑 33(3),248
453)河野太郎(1967) 全国蚕種研究会第12回研修会記録,5-48
454)河野太郎,小川克明(1965) 日蚕雑 34(3),203-204
455)河野太郎,小川克明(1967) 日蚕雑 36(3),253
456)*久保田正樹(1938) 蚕業評論 21(5)
457)工藤政市,徳重直衛(1925) 大日本蚕糸会報 34(399),383-386
458)功刀文良(1958) 蚕糸研究 (27),1-3
459)倉沢一二三,河野太郎,窪田 厚,小川克明(1966) 日蚕雑 35(3),222
460)倉田啓而(1967) 日蚕雑 36(5),436-437
461)倉田啓而,高橋澄雄(1967) 日蚕雄 36(3),235
462)倉田和平,四方栄市(1956) 蚕糸研究 (16),1-2
463)栗栖弌彦(1962) 日蚕雑 31(3),193-194
464)栗栖弌彦(1963) 日蚕雑 32(3),178
465)栗栖弌彦(1965) 京都工芸繊雄大繊維報 4(3),285-290
466)*黒田嘉一郎(1934) 朝鮮化学会報 5(10)
467)黒田嘉一郎,李基寧(1943) 植物及動物 11(6),467-470
468)黒沢益雄(1934) 衣笠蚕報 (336)、86-105
469)来海 栄,門脇 浩(1955) 島根蚕試報 (33),8-11
470)楠野正夫(1956) 日蚕雄 25(6),422
471)沓掛久雄(1940) 蚕試彙 (57),33-53
472)沓掛久雄(1944) 日蚕雑 15(1・2・3・4),18-29
473)沓掛久雄(1952) 鐘紡蚕研報,1-89
474)沓掛久雄(1954) 日蚕東海 (2),36-40
475)沓掛久雄(1961) 全国蚕種協会講習会資料
476)沓掛久雄,黒岩久平(1949) 技術資料 (11),11
477)沓掛久雄,黒岩久平(1951) 技術資料 (29),19-20
478)沓掛久雄,黒岩久平(1952) 日蚕雑 21(4),163-166
479)沓掛久雄,黒岩久平(1963) 日蚕雑 32(3),191-192
480)沓掛久雄,黒岩久平,正木卓治(1961) 日蚕雑 30(3),248-249
481)沓掛久雄,長島栄一(1963) 日蚕雑 32(3),192
482)沓掛克己(1949) 日蚕雑 18(5),325
483)桑名寿一,石井五郎,中村茂子外3名(1964) 蚕糸研究 (52),44-57
484)桑名寿一,高見丈夫(1957)無脊椎動物発生学(久米叉三,団勝磨編)第10章Ⅲ.培風館.東京
485)桑野恒雄(1950) 日蚕雑 19(5),464-468
486)桑野恒雄(1952) 日蚕維 21(2・3),101-105
487)桑野恒雄(1955) 日蚕雑 24(3),145-149
488)桑田和子,山本恵美子,浜村保次(1968) 日蚕雑 37(1),63-65
489)町田二郎(1921) 佐久良会誌 (10),1-5
490)町田二郎(1922) 蚕試報 6(2),35-80
491)町田二郎(1922) 蚕試彙 (16),1-14
492)町田二郎(1924) 蚕業新報 32(367),36-40
493)町田二郎(1929) 応動 1(1),9-13
494)町田二郎(1930) 蚕糸界報 (455),66-70
495)町田二郎(1935) 動雑 47(562・3),521-534
496)町田二郎(1940) 蚕試報 10(1),27-67
497)町田二郎,高梨亮次郎(1929) 佐久良会誌 (24),1-6
498)町田二郎,渡辺勘次(1927) 蚕業新報 35(406),454-472
499)MACHIDA,J.(1920) Zeit.Zell mikr.Anat. 9(3),466-510
500)MACHIDA,J.(1926) Jour.Coll.Agr.Imp.Univ.Tokyo 7(4),293-351
501)MACHIDA,J.(1941) Jour.Coll.Agr.Imp.Univ.Tokyo 15(1),83-131
502)前田富八(1933) 衣笠蚕報 (325),213-225
503)松原藤好(1965) 京都工芸繊雄大繊維報 4(3),291-297
504)松原藤好(1966) 京都工芸繊雄大繊維報 5(1),1-6
505)松原藤好(1967) 日蚕雑 36(3),151-158
506)松原藤好(1967) 日蚕雑 36(3),159-164
507)松原藤好(1968) 日蚕雑 37(2),137-143
508)松原藤好,林屋慶三,野村和子(1964) 日蚕雑 33(3),243-244
509)松田正人,石井博安(1962) 日蚕雑 31(3),183
510)松田基一,樋口芳吉,須藤光正外5名(1966) 日蚕雑 35(4),285-288
511)松井久二郎(1933) 衣笠蚕報 (317),1-15 ; (318),23-32
512)松村季美(1928) 長野蚕試報 (6),1-54
513)松村季美(1939) 蚕糸雑 11(3),164-169
514)松村季美,樋口琢磨,北島正生(1928) 長野蚕試報 (4),1-80
515)松村季美,石坂義高(1929) 長野蚕試報 (9),1-136
516)松村季美,石坂義高(1930) 長野蚕試報 (10),1-40
517)松村季美,竹内好武(1950) 日蚕雑 19(2),192-200
518)松村季美,田中茂明(1952) 技術資料 (33),32
519)松村季美,田中茂明,高坂孝義(1955) 蚕試彙 (69),1-47
520)松村季美,和田盛直(1953) 蚕糸研究 (3),48-51
521)松室重正(1916) 蚕業新報 24(279),7-13
522)松永信義(1934) 朝鮮総督府農試蚕糸部報 3(3),85-143
523)松尾寛一,立川松男,後藤哲翁(1956) 日蚕九州 (昭31),24
524)松崎 巌(1952) 日蚕東北 (5),38
524a)松崎守夫(1968) 日蚕雑 37(6),483-490
525)日黒忠義(1943) 新潟蚕試彙 (4),21-28
526)日黒忠義(1949) 日蚕雑 18(3),169-170
527)緑川栄一,小林悦雄,阿相敏雄(1963) 日蚕雑 32(3),171
528)緑川栄一,近藤 実(1956) 蚕研彙 (6),1-22
529)三重県原蚕種製造所(1922) 三重原蚕成績 (5),53-55
530)三重県蚕業試験場(1923) 三重蚕試成績 (6),30-36
531)三重県蚕業試験場(1923) 三重蚕試成績 (6),36-38
532)見波定治(1922) 農学会報 (234),189-223
533)蓑谷香苗(1926) 埼玉蚕試報 (13),1-45
534)蓑谷香苗(1926) 大日本蚕糸会報 35(418),1377-1387
535)蓑谷香苗(1928) 埼玉蚕試報 (17),1-75
536)蓑谷香苗(1931) 日蚕雑 2(2),186-187
537)蓑谷香苗(1932) 日蚕雑 3(1),70-86
538)蓑谷香苗(1932) 蚕糸界報 41(490),48-54
539)三谷賢三郎(1948) 日蚕雑 17(1・2),40-44
540)三谷賢三郎(1957) 蚕糸界報 66(781),31-34
541)三谷賢三郎,伊与田茂,伊藤節之,梅村角一(1930) 愛知蚕試報 (2),108-152
542)三谷賢三郎,金井 保(1950)日蚕雑 19(1),40-45
543)三谷賢三郎,金井 保(1950) 日蚕雑 19(5),469474
544)三谷賢三郎,金井 保(1951) 日蚕雑 20(5),370-372
545)三谷賢三郎,渡会寿太郎(1916) 愛知県蚕報 (3),120-130
546)三浦英太郎(1923) 衣笠蚕報 (202),29-47
547)三浦英太郎(1925) 人工孵化蚕種学. 中島書院,京都
548)三浦英太郎(1932) 京都高蚕報 1(3),1-36
549)三浦英太郎(1932) 京都高蚕報 1(3),37-42
550)三浦英太郎(1938) 蚕糸界報 47(559),35-39
551)三浦英太郎(1938) 京都高蚕報 2(1),1-56
552)*三浦英太郎(1939) 蚕糸情報 2(1),17-21
553)MIURA,E.(1929) Bull.Imp.Kyoto Sericult.Coll. 1(2),1-44
554)宮慶一郎(1959) 日蚕雑 28(3),163-164
555)宮慶一郎(1959) 日蚕東北 (13),17
556)宮慶一郎(1960) 日蚕雑 29(3),273
557)宮慶一郎(1960) 日蚕東北 (14),43
558)宮慶一郎,沼田六七八(1961) 日蚕雑 30(3),249
559)宮慶一郎,沼田六七八(1962) 日蚕雑 31(3),190
560)MIYA,K.(1956) Jour.Fac.Agr.Iwate Univ. 2(4),393-400
561)MIYA,K.(1958) Jour.Fac.Agr.Iwate Univ. 3(4),436-467
562)MIYA,K.(1959) Jour.Fac.Agr.Iwate Univ. 4(2),126-151
563)MIYA,K.and KURIHARA,M.(1966) Jour.Fac.Agr.Iwate Univ. 8(1),1-13
564)宮川千三郎(1952) 蚕糸研究 (2),40-44
565)宮川千三郎,一場静夫(1965) 蚕糸研究 (55),45-49
566)宮川千三郎,小岩 □(1962) 蚕糸研究 (42),1-5
567)三宅康次,田所哲太郎(1911) 大日本蚕糸会報 (239),。1-4
568)宮坂 収(1935) 蚕糸雑 8(1・2),117-126
569)宮崎重美(1927) 中央蚕糸報 (132),74-76
570)宮崎県原蚕種製造所(1921) 宮崎県原蚕報 (1),212-220
571)宮沢 誉(1951) 技術資料 (29),20
572)宮沢 誉(1954) 日蚕雑 23(3),182
573)水井寿一郎.稲熊真市(1910) 東京蚕講報 (40),141-172
574)水野 保,浅羽登代夫,高野 平,飯村重男(1933) 茨城蚕試彙 (3),1-33
575)水野辰五郎(1920) 蚕試報 4(4),205-262
576)水野辰五郎(1920) 佐久良会誌 (7),32-39
577)水野辰五郎(1920) 佐久良会誌 (8),41-43
578)水野辰五郎(1921) 佐久良会誌 (10),57-60
579)水野辰五郎(1922) 佐久良会誌 (11),28-34
580)水野辰五郎(1929) 蚕卵論.明文堂,東京
581)水野辰五郎,平田四郎右衛門(1927) 佐久良会誌 (21),20-26
582)水田美照(1947) 日蚕東北 (1),36-37
583)水田美照(1949) 日蚕東北 (2),44-47
584)水田美照(1951) 日蚕東北 (4),49-51
585)水田美照,佐藤 誠(1963) 蚕糸研究(47),13-25
586)水田美照,佐藤 誠(1964) 日蚕雑 33(1),59-62
587)水田美照,高橋澄雄(1958) 日蚕雑 27(3),104-106
588)水田美照,高橋澄雄(1958) 日蚕雑 27(4),247-252
589)水田美照,高橋澄雄,苫米地貞夫(1960) 蚕試彙 (76),25-43
590)水田美照,苫米地貞夫(1962) 日蚕雑 31(5),355-358
591)文部省(著作権所有者)(1963) 蚕種製造.実数出版株式会社.東京
592)森本 彰(1956) 日蚕関西 (13),22
593)盛次 清(1937) 衣笠蚕報 (371),118-123
594)盛次 清(1938) 衣笠蚕報 (378),42-52
595)MORIYAMA,T.(1935) 衣笠蚕報 (345),80-84
596)諸星静次郎(編)(1956) 蚕糸界報 65(760),34-38;(762),34-41;(763),29-34;(764),26-32
597)諸星静次郎(1959) 実験形態学新説(竹脇 潔,針塚正樹,深谷昌次編)第3編.養賢堂,東京
598)諸星静次郎(1967) 全国蚕種研究会第12回研修会記録,49-95
599)諸星静次郎,小林宜夫(1949) 日蚕雑 18(5),332-333
600)村上昭雄(1967) 日蚕雑 36(3),254-255
601)村上昭雄,鬼丸喜美治(1966) 日蚕雑 35(3),223
602)MURAKAMI,A.(1966) Ann.Rept.Nat.Inst.Gen.Japan (16),107-109
603)室賀兵左衛門(1926) 蚕業評論 9(5),25-28
604)室賀兵左衛門(1930) 応動 2(1),5-10
605)室賀兵左衛門(1934) 蚕業新報 42(1),37-41
606)室賀兵左衛門(1934) 応動 6(2),84-88
607)室賀兵左衛門(1934) 蚕糸界報 43(509),79
608)室賀兵左衛門(1935) 日蚕雑 6(1),1-5
609)室賀兵左衛門(1935) 日蚕雑 6(1),6-8
610)室賀兵左衛門(1935) 日蚕雑 6(3),175-183
611)室賀兵左衛門(1939) 蚕糸雑 11(4),258-264
612)室賀兵左衛門(1939) 日蚕雑 10(3),150-155
613)室賀兵左衛門(1940) 応動 12(1),13-20
614)室賀兵左衛門(1941) 応動13(1),39-48
615)室賀兵左衛門(1941) 日蚕雑 12(3),136-142
616)室賀兵左衛門(1943) 日蚕雑 14(5・6),237-249
617)室賀兵左衛門(1943) 日蚕雑 14(5・6),250-262
618)室賀兵左衛門(1947) 日蚕雑 16(1・2),20-25
619)室賀兵左衛門(1949) 日蚕雑 18(2),120-121
620)室賀兵左衛門(1949) 新らしい養蚕の原理. 泰文館,東京
621)室賀兵左衛門(1951) 日蚕雑 20(2),92-94
622)室賀兵左衛門(1958) 蚕種学要綱.大推堂,京都,
623)室賀兵左衛門,丸山徳雄(1929) 蚕糸界報 (448),719-721
624)室賀兵左衛門,太田安澄,柏田 豊外3名(1951) 日蚕雑 20(2),161
624a)長江三木夫(1963) 徳島蚕試蚕糸技術 10(1),12
625)永井 覚(1934) 郡是蚕事所彙 (1),57-73
626)永井 覚(1934) 郡是蚕事所彙 (1),75-92
627)永井 覚(1935) 郡是蚕事所彙 (2),45-52
628)永井 覚(1935) 郡是蚕事所彙 (2),53-60
629)永井 覚(1935) 郡是蚕事所彙 (2),61-73
630)永井 覚(1936) 応動 8(1),28-37
631)永井 覚(1941) 日蚕雑 12(3),232-234
631a)永井 覚(1950) 日蚕雑 19(2),126-135
632)長池幸男(1961) 日蚕九州 (昭35),14
633)長池幸男(1961) 日蚕九州 (昭35),14
634)長野県蚕業取締所飯田支所(1934) 天竜蚕糸 7(74),57-58
635)長島栄一(1956) 日蚕雑 25(4),288-294
636)長島栄一,富田克衛,清水隆三(1960) 日蚕臨時大会講演集(中部),13-14
637)長田貞一(1951) 日蚕雑 20(5),335-337
638)長田貞一(1955) 日蚕関西 (11),9-10
639)長田貞一(1960) 蚕糸界報 69(816),27-33
640)長田貞一(1960) 衣笠学報 (8),13-15
641)長田貞一(1960) 衣笠学報 (8),16-18
642)長田貞一(1960) 医学と生物学 57(3),90-93
643)長田貞一(1960) 医学と生物学 57(3),110-113
644)長田貞一(1963) 蚕卵簡易解剖実験指針(学生実験用プリント)
645)永友雄(1931) 日蚕雑 2(2),203-204
646)永友雄(1942) 日蚕雑 13(3 ),114-115
647)永友雄(1943) 日蚕雑 14,201
648)永友雄(1949) 日蚕雑 18(5),330-331
649)中井五二(1925) 実験蚕卵稀塩酸人工孵化法.明文堂,東京
650)中島晴海(1960) 日蚕東北 (14),20-21
651)中島晴海,佐藤喜美雄(1961) 日蚕東北 (15),27-28
652)中島 誠,田島弥太郎(1957) 日蚕雑 26(3),235
653)中島 茂(1938) 蚕糸雑 11(1),56-59
654)中村島次郎,日高末吉(1929) 蚕糸界報 (447),589-594
655)中西邦男(1954) 日蚕東海 (2),22
656)仲野良男(1933) 群馬蚕試報 (16),1-38
657)仲野良男(1933) 群馬蚕試報 (17),1-83
658)仲野良男(1935) 群馬蚕試報 (21),1-135
659)仲野良男(1935) 群馬蚕試報 (22),45-78
660)仲野良男(1951) 日蚕雑 20(4),232-248
661)仲野良男,北田久夫(1951)日蚕東北 (4),44-46
662)中曽根長男(1930) 蚕糸雑 2(1),71-77
663)中曽根長男(1938) 日蚕雑 9(3),310-312
664)中曽根長男(1938) 蚕糸雑 11(2),69-79
665)中曽根長男(1939) 日農化誌 15(10),982-988
666)中曽根正一(1965) 日蚕雑 34(5),333-337
667)中田太郎(1927) 九大農学芸 2(5),399-404
668)中田太郎(1932) 福岡蚕試報 1(2),1-26
669)中田太郎(1936) 日蚕雑 7(3),263-265
670)中田太郎(1952) 郡是研究所彙 (9),1-88
671)中田太郎,宮沢 誉(1950) 日蚕雑 19(5),481-489
672)中田太郎,宮沢 誉(1951) 日蚕雑 20(4),285-286
673)中山吉二,松田 浩(1934) 沖縄蚕試報 (3),85-127
674)中山正幸(1958) 蚕糸研究 (25),1-6
675)難波重雄(1960) 日蚕東北 (14),12-13
670 西村国男,降旗則子(1958) 日蚕雑27(3),155
677)西村国男,降旗則子,井古田尚子(1959) 日蚕雑 28(1),10-16
678)西村国男,山崎 寿(1958) 日蚕雑 27(1),1-4
679)西躰隆雄,山本 巌(1958) 蚕糸研究 (27),11-15
680)西沢一俊,小林敏雄(1955) 日蚕雑 24(5・6),318-324
681)西沢一俊,清水 猛,大日方仁,山ロ定次郎(1953) 日蚕雑 22(4),143-148
682)西沢正一(1965) 蚕糸科学と技術 4(3),68-71
683)入戸野康彦,長池幸男(1952) 日蚕雑 21(2-3),93-94
684)入戸野康彦,中曽根正一(1956) 日蚕雑 25(5),317-321
685)入戸野康彦,大河内徳洽,難波チヨ子(1961) 蚕試彙 (78),15-22
686)入戸野康彦,竹下弘夫(1953) 日蚕雑 22(3),131
687)入戸野康彦,竹下弘夫(1953) 日蚕雑 22(4),149-154
688)入戸野康彦,竹下弘夫(1954) 日蚕雑 23(1),44-48
689)野口活也(1940) 応動 12(3・4),134-138
690)野口活也(1949) 日蚕東北 (2),47-49
691)野口活也,倉沢恒夫,安藤孝雄(1960) 日蚕東北(14),26
692)野尻邦雄(1965) 蚕糸研究 (54),13-14
693)野尻邦雄,高山善助(1961) 日蚕東北 (15),4-5
694)農林省蚕糸試験場(1954) 蚕試資料 (16),1-12
695)布目順郎(1937) 応動 9(2),68-92
696)布目順郎(1943) 日蚕雑 14(5・6),228-236
697)布目順郎(1950) 日蚕雑 19(4),315-322
698)布目順郎,八幡 穣,赤木八州夫(1955) 京都工芸繊維大繊維報 1(2),177-185
699)小田中久太郎(1936) 熊本蚕試彙 (6),49-65
700)岡田一次,広部達道,下口曠志(1964) 玉川大農報 (5),33-42
701)岡田 清(1960) 日蚕東海 (8),24
702)岡田益吉(1967) 実験形態学誌 (21),495
703)OKADA,M.(1958) 応動昆 2(4),295-296
704)岡村郡重(1926) 佐久良会誌 (19),40-44
705)岡村郡重(1927) 佐久良会誌 (21),97-101
706)岡村郡重(1927) 佐久良会誌 (22),81-83
707)奥村清久(1918) 佐久良会誌(4),50-60
708)小村達夫(1951) 動雑 60(3),68-69’
709)鬼火喜美治(1964) 日蚕雑 33(3),250
710)小野秀記,斎田哲夫(1951) 技術資料 (29),23
711)*小野正武(1942) 鹿児島高農報 (14),123-160
712)小野正武(1942) 蚕試報 10(8),495-542
713)小野正武(1951) 蚕試報 13(6),247-303
714)小野寺秀夫(1950) 日蚕東北 (3),99-101
715)小野寺秀夫(1952) 日蚕東北 (5),26-28
716)小野寺秀夫,鈴木隆七(1951) 日蚕東北 (4),55-56
717)小山内実,波磨忠雄(1962) 動雑 71(11・12),381
718)押部秀二(1937) 衣笠蚕報 (371),158-170
719)尾崎準一(1947) 蚕糸化学と副産物利用.朝倉書店,東京
720)小沢民治(1959) 日蚕雑 28(4),211-219
721)小沢民治(1965) 日蚕雑 34(3),196
722)小沢民治(1966) 日蚕雑 35(3),233
723)大場治男(1942) 植物及動物 10(7),687
724)大場治男(1951) 日蚕雑 20(3),186-187
725)大場治男(1952) 日蚕雑 21(1),28
726)大場治男(1952) 長野蚕試蚕糸技術 15(12),1-4
727)大場治男,中島利広(1954) 長野蚕試蚕糸技術 17(12),283-292
728)大場治男,中島利広(1957) 長野蚕試報 10(6),382-389
729)大江亀久(1939) 蚕糸界報 48(569),40-45
730)大櫛平三郎,地引嘉作(1925) 熊本蚕試報 1(4),395-436
731)大櫛平三郎,地引嘉作,広田利之(1925) 熊本蚕試報 1(3),269-394
732)大櫛平三郎,地引嘉作,広田利之(1925) 熊本蚕試報 1(4),439
445
733)大櫛平三郎,地引嘉作,広田利之(1926) 熊本蚕試報 2(1),45-75
734)大櫛平三郎,地引嘉作,広田利之,桐野義盛(1925) 熊本蚕試報 1(2),89-268
735)大島 格(1960) 応動昆 4(4),212-225
736)大久保紀元(1966) 日蚕雑 35(2),81-84
737)大宮新左衛門(1947) 日蚕東北 (1),57-61
738)大宮新左衛門(1960) 日蚕東北 (14),42-43
739)大宮新左衛門(1961) 蚕糸研究 (39),5-11
740)大宮新左衛門(1961) 蚕糸研究 (39),27-37
741)大宮新左衛門(1964) 蚕糸研究 (49),25-30
742)大宮新左衛門(1964) 蚕糸研究 (52),7-19
743)大宮新左衛門,中島晴海(1966) 蚕糸研究 (59),16-22
744)大宮新左衛門,大河内徳治(1953) 蚕糸研究 (4),11-13
745)大宮新左衛門,滝沢寛三(1957) 蚕糸研究 (20),47-49
746)大村清之助(1935) 日蚕雑 6(2),159-161
747)大村清之助(1937) 日蚕雑 8(2),162
748)大村清之助(1939) 日蚕雑 10(2),49-57
749)大村清之助(1941) 遺雑 17,295-303
750)大村清之助(1942) 日蚕雑 13(2),30-38
751)大村清之助(1942) 日蚕雑 13(2),39-48
752)大村清之助(1947) 日蚕東北 (1),31-32
753)大村清之助(1947) 日蚕東北 (1),32-34
754)大村清之助(1949) 日蚕雑 18(5),314-315
755)大村清之助(1950) 蚕試報 13(3),79-130
756)大村清之助,片岡 平(1943) 日蚕雑 14(5・6),263-275
757)大村清之助,菅生文雄(1949) 日蚕東北 (2),50-51
758)OMURA,S.(1936) Jour.Fac.Agr.Hokkaido Imp.Univ. 38(2),151-181
759)OMURA,S.(1938) Jour.Fac.Agr.Hokkaido Imp.Univ. 40(3),111-128
760)OMURA,S.(1938) Jour.Fac.Agr.Hokkaido Imp.Univ. 40(3),129-170
761)大西重信(1954) 宮崎蚕試報 (5),1-6
762)大西重信(1954) 宮崎蚕試報 (5),6-15
763)大西重信,高橋尚徳(1961) 日蚕九州 (昭35),20
764)大野稜太郎(1949) 技術資料 (13),30-31
765)大野稜太郎(1950) 動雑 59(11),260-263
766)大野稜太郎(1951) 日蚕雑 20(4),297
767)大野稜太郎(1952) 日蚕雑21(2・3),125
768)大貫 道,難波重雄,藤森胡友 外3名(1963) 蚕試彙 (81),5-21
769)大貫 道,押川勝二,長池幸男(1965) 蚕糸研究 (55),1-5
770)大貫 道,押川勝二,中水流操,中村泰郎(1961) 日蚕九州 (昭35),20-21
771)大貫 道,押川勝二,中水流操,中村泰郎(1961) 日蚕九州(昭36),17
772)大沢孝三(1917) 佐久良会誌 (2),79-88
773)大沢孝三(1924) 佐久良会誌 (14),21-30
774)大槻良樹(1965) 京都工芸繊維大繊維報 4(3),314-344
774a)大槻良樹,村上昭雄(1968) 動雑 77(12),383-387
775)大和田亮吉(1924) 佐久良会誌 (16),3-7
776)佐渡敏彦(1960) 遺伝研年報 (10),95
777)佐渡敏彦(1961) 日蚕雑 30(3),249-250
778)佐渡敏彦(1966) 日蚕雑 35(3),226-227
779)SADO,T.(1961) Jap.Jour.Gen. 36 Suppl. 136-151
780)西郷 力(1936) 日蚕雑 7(4),291-300
781)西郷 力(1938) 日蚕雑 9(3),288-289
782)西郷 力(1951) 技術資料 (29),22
783)西郷 力(1956) 日蚕九州 (昭31),24-25
784)西郷 力(1959) 日蚕九州 (昭34),32
785)西郷 力(1961) 日蚕九州 (昭35),21
786)西郷 力(1961) 日蚕九州 (昭36),18
787)西郷 力(1965) 日蚕雑 34(3),143-148
788)西郷 力(1965) 日蚕雑 34(3),149-154
789)斎藤 広(1960) 日蚕東北 (14),31-32
790)斎藤菊雄(1931) 長野蚕試報 (15),1-26
791)酒井 績,石阪一義(1954) 日蚕中部 (8),1
792)坂之下旭(1967) 九大農学位論文
793)坂手 栄,川口和夫,佐藤清夫(1952) 日蚕雑 21(5-6),237-239
794)崎山用佑(1935) 蚕業新報 43(510),28-29
795)桜井 基,松本 正(1955) 日蚕雑 24(5・6),377-382
796)佐野利男(1965) 静岡蚕試報 (16),21-23
797)蚕種研究室長打合会(1960) 蚕糸研究 (34),14-18
798)蚕種研究室長打合会(1963) 蚕糸研究 (47),9-12
799)蚕種原蚕種研究室長打合会(1962) 蚕糸研究 (43),54-59
800)蚕種原蚕種研究室長打合会(1964) 蚕糸研究 (49),53-57
801)蚕種原蚕種研究室長打合会(1967) 蚕糸研究 (65),74-88
802)蚕種研究室長・原蚕種研究室長打合会(1960) 蚕糸研究 (35),6-17
803)佐々木静(1948) 日蚕雑 17(34),70-74
804)佐々木静(1949) 日蚕雑 18(3),144-148
805)佐々木静(1949) 日蚕雑 18(3),149-151
806)佐々木静(1950) 日蚕雑 19(6),541-547
807)佐々木静(1951) 技術資料 (29),21
808)佐々木静,大槻松夫(1951) 日蚕雑 20(5),346-350
809)佐々木周郁,渡辺忠雄,近藤義和(1957) 日蚕雑 26(4),291-297
810)佐々木太郎(1961) 日蚕東北 (15),6
811)佐瀬 旭(1925) 蚕業新報 33(383),540-542
812)佐藤春太郎(1924) 大日本蚕糸会報 33(383),27-32
; (384),110-114;(386),242-247 ; (387),335-337
813)佐藤春太郎(1925) 農学会報 (273),155-164
814)佐藤春太郎(1926) 農学会報 (284),277-303
815)佐藤春太郎(1934) 応動,6,179-187 ; 225-253
816)佐藤春太郎(1942) 蚕糸雑 14(1),19-27
817)佐藤春太郎(1953) 日蚕雑 22(3),139-140
818)佐藤春太郎,茅野 功(1935) 蚕糸雑 8(1・2),162-179
819)佐藤春太郎,目崎正夫(1942) 蚕糸雑 14(1),28-35
820)佐藤春太郎,田中茂光(1951) 日蚕雑 20(6),418-422
821)SATO,H.(1925) 農学会報 (274),232-238
822)佐藤 広(1947) 日蚕東北 (1),37-40
823)佐藤 広(1952) 日蚕東北 (5),23-25
824)佐藤 広,中島晴海,佐藤喜美雄(1964) 蚕糸研究 (49),58-62
825)佐藤 広,難波重雄,小野寺ナミ子(1961) 日蚕東北 (15),26-27
826)佐藤 広,難波重雄,小野寺ナミ子(1962) 蚕糸研究 (43),43-46
827)佐藤 広,難波重雄,佐藤喜美雄(1964) 日蚕雑 33(1),72-76,
828)佐藤 広,高見丈夫,北沢敏男(1956) 日蚕雑 25(4),273-278
829)佐藤市兵衛,杉浦勇治郎(1934) 日蚕雑 5(3),228-231
830)佐藤喜美雄,笹原重雄,佐藤 広(196) 蚕糸研究 (57),13-14
831)佐藤貞治(1936) 日蚕雑 7(1),10-16
832)佐藤貞治(1937) 日蚕雑 8(2),157
833)関 甫(1941) 蚕糸日本 13(11),12-15
834)関戸一正(1950) 日蚕雑 19(6),517-522
835)四方正義(1951) 日蚕雑 20(4),295
836)四方正義(1964) 日蚕雑 33(5),419-423
837)四方正義,吉木聿二(1965) 日蚕雑 34(1),37-41
838)島貫英二(1966) 蚕糸研究 (60),7-13
839)清水隆三,長島栄一(1961) 日蚕雑 30(3),251
840)清水 滋(1929) 応動 1(2),109-116
841)清水 滋(1929) 応動 1(3・4),237-240
842)清水 滋(1935) 応動 7(6),310-312
843)清水 滋(1936) 応動 8(6),315-317
844)清水 滋(1959) 全国蚕種協会講習会資料
845)清水 滋(1961) 日蚕雑 30(3),288-289
846)清水 滋,橋口寿夫(1953) 蚕糸研究 (3),39-41
847)清水 滋,橋口寿夫(1955) 蚕糸研究 (11),27-29
848)清水 滋,橋口寿夫(1956) 蚕糸研究 (15),7-9
849)清水 滋,橋口寿夫(1957) 蚕糸破究 (19),15-18
850)清水 滋,波島千恵子,安江 昇(1966) 蚕糸研究 (58),8-11
851)清水 滋,堀内彬明(1949) 日蚕雑 18(2),95-99
852)清水 滋,堀内彬明(1950) 日蚕雑 19(1),7-15
853)清水 滋,堀内彬明(1950) 日蚕雑 19(2),152-153
854)清水 滋,堀内彬明(1959) 日蚕雑 28(5),277-280
855)清水 滋,堀内彬明(1960) 日蚕雑 29(1),50-51
856)清水 滋,堀内彬明(1962) 日蚕雑 31(1),49
857)清水 滋,堀内彬明,波島千恵子,安江 昇(1962) 日蚕雑 31(3),191
858)清水 滋,堀内彬明,波島千恵子,安江 昇(1963) 日蚕雑 32(2),113-115
859)清水 滋,堀内彬明,昆野民夫(1958) 蚕糸研究 (24),10-12
860)清水 滋,堀内彬明,中村元三,小野寺道子(1960) 蚕糸研究 (33),9-11
861)清水 滋,坂手 栄,中曽根正一(1965) 日蚕雑 34(1),32-36
862)清水 滋,高見丈夫(1955) 蚕試彙 (68),1-37
863)須田金之助(1913) 蚕業新報 21(238),15-18
864)須貝悦治(1963) 日蚕雑 32(3),199-200
865)須貝悦治(1963) 応動昆 7(3),232-233
866)須貝悦治(1963) 応動昆 7(4),353-355
867)須貝悦治(1964) 動雑 73(8),219-222
868)須貝悦治(1965) 動雑 74(9),276-282
869)須貝悦治(1966) 動雑 75(2),ミ4-39
870)須貝悦治,木口憲爾(1967) 日蚕雑 36(6),491-496
871)SUGAI,E. and ASHOUSH,I.(1968) Appl.Ent.Zool. 3(3),99-102
872)杉山八郎(1963) 蚕試彙 (83),1-8
873)杉山八郎,神田俊男(1963) 蚕糸研究 (47),30-36
874)杉山八郎,清水さかき,高見丈夫(1961) 蚕糸研究 (39),16-26
875)杉山八郎,高見丈夫(1959) 蚕糸研究 (32),18-21
876)杉山八郎,高見丈夫(1961) 蚕糸研究 (37),15-23
877)杉山八郎,高見丈夫(1961) 蚕試彙 (78),23-30
878)杉山八郎,高見丈夫(1962) 蚕糸研究 (43),47-53
879)杉山八郎,戸谷和夫(1960) 蚕糸研究 (35),21-22
880)菅生文雄(1955) 群蚕要報 (27),14-16
881)角田文夫,西垣澄敏(1963) 日蚕関西 (26),21
882)角田文夫,西垣澄敏(1963) 日蚕関西 (27),34
883)須藤光正,樋口芳吉,松田基一,外5名(1966) 日蚕雑 35(4),281-284
884)鈴木親?(1959) 全国蚕種研究会講習会記録 (4),55-70
885)鈴木簡一郎(1950) 日蚕雑 19(6),560-570
886)鈴木簡一郎(1951) 蚕試報 13(8),367-385
887)鈴木簡一郎,藤島一民(1952) 技術資料 (33),75-76
888)鈴木簡一郎,平田四郎右衛門(1924) 蚕試報 6(8),301-318
889)鈴木簡一郎,平田四郎右衛門(1925) 佐久良会誌 (17),52-63
890)鈴木簡一郎,大村清之助(1951) 蚕試報 13(8),347-366
891)鈴木簡一郎,辻田光雄(1937) 日蚕雑 8(2),89-105
892)鈴木 清,鈴木親?(1960) 蚕糸研究 (36),1-2
893)鈴木広吉(1917) 蚕試報 2(4),287-312
894)鈴木正親(1960) 日蚕東北 (14),38
895)鈴木 実,鎌倉今朝五郎(1961) 静岡蚕試報 (12),39-46
896)鈴木のぶ(1966) 日蚕雑 35(3),222
897)鈴木のぶ(1967) 日蚕雑 36(3),254
898)鈴木健弘,山桝義寛(1929) 中央蚕糸報 (152),301-308
899)鈴木健弘,山桝義寛(1929) 蚕糸界報 (446),465-470
900)鈴木健弘,山桝義寛,先崎源谷外3名(1929) 衣笠蚕報 (270),1-28
901)鈴木健弘,山桝義寛,先崎源谷外3名(1929) 衣笠蚕報 (271),29-50
902)鈴木健弘,山桝義寛,先崎源谷外3名(1929) 衣笠蚕報 (272),51-74
903)鈴木健弘,山桝義寛,先崎源谷外3名(1929) 衣笠蚕報 (274),75-98
904)高橋伊勢次郎(1912) 大日本蚕糸会報 (246),8-12
905)高橋伊勢次郎,杉田吉重(1912) 東京蚕講成績 (44),63-89
906)高橋幸吉(1932) 日蚕雑 3(3),211-215
907)高橋幸吉(1949) 日蚕雑 18(4),220-226
908)高橋幸吉(1956) 蚕糸会報 65(761),24-29
909)高橋幸吉,中村鶴松,栖栖弌彦外2名(1957) 熊本蚕試報 6(5),1-40
910)高橋清七,田角叉十郎,長谷川正雄(1925) 蚕業新報 33(379),47-54
911)高橋清七,八木誠政(1926) 蚕業新報 (393),266-268
912)高橋保雄(1954) 日蚕雑 23(3),183-184
913)高橋保雄(1955) 長野蚕試蚕糸技術 18(4),93-94
914)高橋保雄(1955) 日蚕雑 24(3),185-186
915)高橋保雄(1956) 日蚕雑 25(3),246
916)高橋保雄(1959) 応動長 3(2),80-85
917)高橋保雄(1960) 日蚕雑 29(3),315
918)高見丈夫(1942) 動雑 54(9),337-343
919)高見丈夫(1943) 動雑 55(6),220-223
920)高見丈夫(1944) 動雑 56(4・5・6),62-65
921)高見丈夫(1944) 動雑 56(4・5・6),66-69
922)高見丈夫(1947)生物 1(4),208-211
923)高見丈夫(1952) 日蚕雑 21(2・3),84-89
924)高見丈夫(1952) 日蚕雑 21(2・3),125
925)高見丈夫(1952) 蚕糸研究 (2),32-37
926)高見丈夫(1952) 日蚕雑 21(5・6),245-251
926a)高見丈夫(1953) 日蚕雑 22(2),58-61
927)高見丈夫(1953) 日蚕雑 22(3),120-121
928)高見丈夫(1953) 日蚕雑 22(5),181-184
929)高見丈夫(1955) 日蚕雑 24(3),184
930)高見丈夫(1957) 蚕試報 14(12),577-594
931)高見丈夫(1959) 蚕試報 15(9),477-492
932)高見丈夫,北沢敏男(1953) 蚕糸研究 (4),17-21
933)高見丈夫,北沢敏男(1959) 蚕糸研究 (31),25-28
934)高見丈夫,北沢敏男(1959) 蚕糸研究 (32),11-17
935)高見丈夫,北沢敏男(1960) 蚕試彙 (75),1-31
936)高見丈夫,北沢敏男(1961) 蚕試彙 (78),1-14
937)高見丈夫,野口和子(1955) 日蚕雑 24(4),254-258
938)高見丈夫,杉山八郎(1964) 日蚕雑 33(3),247
939)高見丈夫,杉山八郎,北沢敏男,神田俊男(1965) 日蚕関東 (16),56-57
940)高見丈夫,杉山八郎,北沢敏男,神田俊男(1966) 蚕糸研究 (58),1-7
941)高見丈夫,杉山八郎,北沢敏男,神田俊男(1966) 応動昆 10(4),197-201
942)高見丈夫,杉山八郎,北沢敏男,神田俊男(1967) 応動昆 11(4),182-186
943)高見丈夫,戸谷和夫,杉山八郎 北沢敏男(1962) 蚕試彙 (79),1-10
944)高見丈夫,戸谷和夫,杉山八郎 北沢敏男(1965) 蚕糸研究 (55),22-26
945)高見丈夫,戸谷和夫,杉山八郎 北沢敏男,神田俊男(1966) 蚕試報 20(2),57-69
946)TAKAMI,T.(1954) Science 119(3083),161-162
947)TAKAMI,T.(1954) Cytologia 19 (4),299-305
948)TAKAMI,T.(1955) Exp.Cell Res. 9 (3),568-571
949)TAKAMI,T.(1958) J.Exp.Biol. 35 (2),286-296
950)TAKAMI,T.(1959) Science 130(3367),98-99
951)TAKAMI,T.(1963) H.Exp.Biol. 40 (4),735-739
952)高梨亮次郎(1927) 大日本蚕糸会報 36(430),1388-1398
953)高梨亮次郎(1927) 佐久良会誌 (21),46-52
954)高梨亮次郎(1946) 蚕卵の保護に就いて.日本蚕糸業会蚕種部
955)高梨亮次郎,足立 潔(1928) 佐久良会誌 (23),10-12
956)高梨亮次郎,松尾重晴(1941) 日蚕雑 12(3),247-248
957)高梨亮次郎,松尾重晴(1944) 日蚕雑 15(1・2・3・4),43-50
958)高岡 実(1967) 動雑 76(11・12),401
959)高崎恒雄,荒武義信(1952) 技術資料 (33),79
960)高崎恒雄,一丸 学(1958) 日蚕雑 27(3),157
961)高瀬慶作(1917) 実験外国種及交雑種蚕児飼育法.明文堂,東京
962)高島成之(1957) 日蚕雑 26(2),53-54
963)高島成之,神田俊男(1963) 蚕糸研究 (47),5-6
964)高島成之,高野幸治(1955) 日蚕雑 24(3),184
965)高島成之,高野幸治(1962) 日蚕雑 31(6),391-397
966)高島成之,高野幸治,神田俊男(1963) 蚕糸研究 (47),7-8
967)高須兵司(未発表の遺稿) 蚕卵の性状調査,蚕卵の破裂抗力に関する研究
968)高取正道(1951) 技術資料 (29),22
969)竹田 寛(1948) 日蚕雑 17(3・4),118-119
970)竹田 寛(1949) 日蚕雑 18(5),335-336
971)竹田 寛(1953) 日蚕雑 22(3),124-125
972)竹田 寛(1953) 日蚕雑 22(3),125
973)竹田 寛(1955) 日蚕雑 24(3),201
974)竹田 寛(1956) 日蚕雑 25(3),246
975)竹田 寛(1957) 日蚕雑 26(3),229
976)竹田 寛(1957) Jour.Fac.Text.Sericult.Shinshu Univ. (13),Ser.A.Biol. (7),1-130
977)竹田 寛,荒井五助,青柳武夫(1967) 日蚕雑 36(3),249
978)竹田 寛,田中一行(1951) 日蚕雑 20(1),74
979)竹田 寛,田中一行(1952) 日蚕雑 21(2・3),127-128
980)竹田 寛,田中一行(1958) 日蚕雑 27(3),171
981)竹田 寛,田中一行(1959) 日蚕雑 28(3),171
982)竹森金次郎(1926) 佐久良会誌 (20),76-78
983)竹内孝三(1955) 日蚕雑 24(1),49-56
984)竹内孝三(1955) 日蚕雑 24(4),259-263
985)竹内孝三(1956) 蚕糸研究 (18),12-15
986)竹内孝三(1957) 蚕糸研究 (19),27-32
987)竹内孝三(1957) 蚕糸研究 (20),25-26
988)竹内孝三(1957) 蚕糸研究 (22),1 -10
989)竹内孝三(1957) 蚕糸研究 (22),11-13
990)竹内孝三(1957) 蚕糸研究 (22),14-18
991)竹内孝三(1958) 蚕糸研究 (25),7 -14
992)竹内孝三(1959) 蚕糸研究 (29),7 -14
993)竹内孝三(1959) 蚕糸研究 (33),5-8
995)竹内重夫(1966) 動雑 75(11・12),329
996)竹内好武,二木猪一,堀内彬明外3名(1960) 蚕試彙 (76),1-24
997)滝沢寛三(1964) 日蚕雑 33(4),355-356
998)滝沢寛三(1967) 日蚕雑 36(1),57-59
999)滝沢寛三,野尻邦雄,八鍬春美(1967) 日蚕雑 36(3),223-227
1000)滝沢寛三,大友昭子(1960) 日蚕東北 (14),36-39
1001)滝沢寛三,大友昭子(1964) 蚕糸研究 (49),39-41
1002)滝沢義郎(1951) 日蚕東北 (4),61-63
1003)滝沢義郎(1951) 技術資料 (29),50
1004)滝沢義郎,平田貞雄(1950) 日蚕東北 (3),94-95
1005)滝沢義郎,勝野貞哉(1959) 日蚕東北 (13),38
1005a)滝沢義郎,勝野貞哉(1961) 日蚕東北 (15),1
1006)滝沢義郎,久保 剛(1952) 日蚕東北 (5),17
1007)滝沢義郎,七戸英三(1950) 日蚕東北 (3),90-92
1008)滝沢義郎,七戸英三(1951) 日蚕雑 20(1),49
1009)滝沢義郎,玉沢 享(1952) 日蚕東北 (5),15-17
1010)滝沢義郎,玉沢 享(1959) 日蚕雑 28(3),170-171
1011)滝沢義郎,玉沢 享(1959) 日東蚕北 (13),24
1012)滝沢義郎,玉沢 享(1960) 日蚕東北 (14),39-41
1013)滝沢義郎,玉沢 享(1961) 日蚕東北 (15),34
1o14)玉沢 享(1966) 日蚕雑 35(3),228
1015)田辺七郎(1950) 技術資料 (22),19-20
1016)田辺七郎(1951) 技術資料 (29),20-21
1017)田辺七郎,萩原博武(1954) 広島蚕試成績 (1),48-50
1018)田辺七郎,萩原博武(1955) 広島蚕試成績 (2),21-23
1019)田中 深(1951) 技術資料 (29),22
1020)田中 深(1964) 日蚕雑 33(3),249
1021)田中 深(1966) 日蚕雑 35(2),88-94
1022)田中 深(1966) 日蚕雑 35(3),165-168
1023)田中 深(1966) 日蚕雑 35(3),210-211
1024)田中 深(1966) 日蚕雑 35(4),262-266
1025)田中 深(1966) 日蚕雑 35(5),321-326
1026)田中 深(1966) 日蚕雑 35(5),327-330
1027)田中 深,中光愛幸(1964) 日蚕雑 33(3),248-249
1028)田中孝平,福沢久雄(1962) 群蚕要報 (48),15-26
1029)田中孝平,神 保義(1955) 群蚕要報 (27),8-14
1030)田中茂光,大井昌次,佐藤春太郎(1955) 信大繊維報 (5),63-67
1031)TANAKA,S.(1961) Jour.Fac.Text.Sericult.Shinshu Univ. (29),Ser.E.Agr.Sericult. (5),69-123
1032)田中茂男(1965) 日蚕雑 34(5),369-370
1033)田中茂男(1967) 日蚕雑 36(3),177-182
1034)田中恒夫(1956) 日蚕雑 25(3),217-218
1035)田中義麿(1928) 蚕体解剖学講義 上巻. 明文堂,東京
1036)田中義麿(1943) 蚕学.興文社,東京
1037)TANAKA,Y.(1953) Advances in Genetics 5, 239-317
1038)谷口幸雄(1959) 日蚕雑 28(3),162-163
1038a)谷口幸雄(1960) 日蚕雑 29(3),274
1039)谷口幸雄(1961) 日蚕雑 30(3),247
1040)田島弥太郎(1938) 遺雑 14,117-128
1041)田島弥太郎(1941) 日蚕雑 12(3),184-188
1042)田島弥太郎(1942) 日蚕雑 13(3),81-94
1043)田島弥太郎(1943) 日蚕雑 14(1・2),76-89
1044)田島弥太郎(1943) 日蚕雑 14(1・2),90-97
1045)田島弥太郎(1944) 蚕試報 12(2),109-181
1046)田島弥太郎(1947) 蚕のモザイク.北陸館,東京
1047)田島弥太郎,原口忠次,太田 登,外2名(1955) 蚕研彙 (5),25-47
1048)田島弥太郎,久米富男,上岡政美(1955) 蚕研彙 (5),5-24
1049)田島弥太郎,緑川栄一,太田 登,小林義彦(1955) 蚕研彙 (5),48-61
1050)田島弥太郎,大沼昭夫(1966) 日蚕雑 35(3),223-224
1051)TAJIMA,Y. and ONIMARU,K.(1965) Ann.Rept.Nat.Inst.Gen.Japan (15),44-46
1052)TAJIMA,Y. and ONUMA,A.(1966) Ann.Rept.Nat.Inst.Gen.Japan (16),38-39
1053)TAJIMA,Y. and ONUMA,A.(1967) 日蚕雑 36(4),286-292
1054)東畑道夫(1924) 蚕糸学報 6(5),25-26
1055)東畑道夫,功力文良(1941) 日蚕雑 12(4),282-289
1056)徳永雅明(1939) 応動 11(3・4),154-159
1057)TOMITA,M.(1921) Biocem.Zeit. 116,40-47
1058)鳥浜義己(1956) 日蚕雑 25(3),206-207
1059)鳥浜義己(1964) 日蚕雑 33(3),243
1060)鳥居一男,森井謙介(1948) 蚕研彙 2(1),2-3
1061)戸谷和夫(1953) 日蚕雑 22(6),248-251
1062)戸谷和夫(1953) 日蚕雑 22(6),251-253
1063)戸谷和夫(1956) 日蚕雑 25(4),268-272
1064)戸谷和夫(1958) 蚕糸研究 (24),7-9
1065)戸谷和夫(1958) 日蚕雑 27(6),400-402
1066)戸谷和夫(1960) 日蚕雑 29(3),273
1067)戸谷和夫(1960) 信大繊維報 (10),118-123
1068)戸谷和夫(1964) 日蚕雑 33(3),248
1069)戸谷和夫(1968) 日蚕雑 37(5),420-426
1070)戸谷和夫,早坂昭二(1967) 日蚕雑 36(5),400-402
1070a)戸谷和夫,中島晴海,苫米地貞夫(1968) 蚕糸研究 (69),24-26
1071)戸谷和夫,清水さかき(1959) 蚕糸研究 (32),7-10
1072)戸谷和夫,清水さかき(1960) 蚕糸研究 (34),1-3
1072a)戸谷和夫,滝沢寛三,苫米地貞夫(1968) 蚕試彙 (91),1-6
1073)外山亀太郎(1896) 農学会報 (28),28-40
1074)外山亀太郎(1909) 蚕種論.丸山舎,東京
1075)TOYAMA,K.(1902) Bull.Agr.Tokyo Imp.Univ.Japan 5 (1),73-118
1076)土屋久一,加藤 久,叶内順治(1952) 技術資料 (33),1-2
1077)土屋久一,富塚喜吉(1951) 日蚕東北 (4),89-90
1078)土屋登美次(1926) 蚕業新報 34(392),155-157
1079)辻陽太郎,藤崎卓爾(1903) 東京蚕講報 (19),30-39
1080)辻陽太郎,谷田部七五郎(1901) 東京蚕講報 (17),67-77
1081)辻田光雄(1948) 蚕試報 12(6),633-648
1082)辻田光雄(1948) 日蚕報 17(3・4),62-69
1083)辻田光雄(1951) 蚕試報 13(8),387-421
1084)辻田光雄,梅谷与七郎(1953) 蚕試報 14(3),93-113
1085)塚越次郎(1935) 蚕糸界報 44(517),80-83
1086)塚越次郎(1957) 蚕糸研究 (22),36-42
1087)堤 要造,藤森胡友,清水文信(1959) 蚕糸研究 (29),21-25
1088)堤 要造,清水文信(1961) 蚕糸研究 (37),28
1089)内山安平,小鉢春利(1952) 山梨蚕試彙 (15),36-42
1090)上田金時(1961) 日蚕雑 30(4),313-318
1091)上原 功(1953) 日蚕雑 22(3),121-122
1092)上原 功(1951) 埼玉蚕試要報 (27),44
1093)上原 功(1955) 日蚕雑 24(3),186
1094)上原 功(1956) 日蚕雑 25(3),219-220
1095)梅原宗一(1963) 日蚕関西 (27),1
1096)梅谷与七郎(1927) 朝鮮総督府勧業模範場蚕試報 2(1),1-33
1097)梅谷与七郎(1929) 日本学術協会報 5,395-403
1098)梅谷与七郎(1935) 動雑 47(562・3),659-671
1099)梅谷与七郎(1937) 日蚕雑 8(4),297-310
1100)梅谷与七郎(1937) 動雑 49(9),300-302
1101)梅谷与七郎(1937) 朝鮮総督府農試蚕糸報 4(1),1-36
1102)梅谷与七郎(1938) 蚕糸界報 47(560),33-38
1103)梅谷与七郎(1939) 日蚕報 10(2),64-76
1104)梅谷与七郎(1939) 日本学術協会報 14(3),504-508
1105)梅谷与七郎(1940) 日蚕雑 11(2),111-112
1106)梅谷与七郎(1946) 蚕試報 12(4),393-480
1107)梅谷与七郎(1948) 日蚕雑 17(3・4),117-118
1108)梅谷与七郎(1949) 動雑 58(6),94-95
1109)梅谷与七郎(1949) 日蚕雑 18(5),267-270
1110)梅谷与七郎(1950) 動雑 59(12),272-277
1111)梅谷与七郎(1950) 動雑 59(12),278-283
1112)梅谷与七郎(1952) 日蚕雑 21(1),53
1113)梅谷与七郎(1954) 文部省科学試験研究費による研究報告集録 農学編,329-330
1114)梅谷与七郎(1955) 遺雑 30(3),133-138
1115)梅谷与七郎(1955) 日蚕雑 24(4),267-272
1116)梅谷与七郎,浅野清志(1952) 動雑 61(3・4),83
1117)梅谷与七郎,大村清之助(1950) 蚕試報 13(2),63-78
1118)梅谷与七郎,大野稜太郎(1949) 日蚕雑 18(5)337-338
1119)梅谷与七郎,大野稜太郎(1950) 日蚕雑 19(2),136-146
1120)梅谷与七郎,戸谷和夫(1949) 日蚕雑 18(5),271-273
1121)梅谷与七郎,渡辺喜二郎(1959) 蚕糸研究 (28),4-5
1122)UMEYA,Y.(1926) Jour.Coll.Agr.Imp.Univ.Tokyo 9(1),57-84
1123)UMEYA,Y.(1937) Proc.Imp.Acad. 13(9),378-380
1124)UMEYA,Y.(1955) Cytologia 20 (3),267-272
1125)梅津富雄(1952) 日蚕東北 (5),39-40
1126)浦城常徳,北田久夫(1953) 福島蚕試彙 (11),90-91
1127)牛込正一(1930) 群馬蚕試報 (8),1-106
1128)牛込正一,服部文雄(1928) 群馬蚕試報 (4),1-143
1129)内海 進,栗栖弌彦,市川吉夫 外3名(1966) 日蚕雑 35(6),405-410
1130)和田盛直(1956) 蚕糸研究 (16),3-5
1131)和田昭治(1954) 日蚕雑 23(6),339-342
1132)渡部 仁(1961) 日蚕雑 30(2),95-100
1133)渡部 仁(1961) 日蚕雑 30(4),345-350
1134)渡辺勘次(1918) 蚕試報 3(8),397-437
1135)渡辺勘次(1918) 蚕試報 3(8),439-456
1136)渡辺勘次(1919) 蚕試報 4(2),87-106
1137)渡辺勘次(1919) 蚕試報 4(2),107-116
1138)渡辺勘次(1922) 蚕試彙 (9),1-15
1139)渡辺勘次(1922) 蚕試彙 (16),1542
1140)渡辺勘次(編)(1924) 蚕試彙 (22),17-54
1141)渡辺勘次(1924) 蚕試報 6(9),411-455
1142)渡辺勘次(1926) 蚕業新報 34(402),1359-1368
1143)渡辺勘次(1927) 佐久良会誌 (21),26-33
1144)渡辺勘次(1928) 蚕試報 7(7),285-303
1145)渡辺勘次(1931) 蚕試彙(41),16-30
1146)渡辺勘次(編)(1934) 蚕試彙 (45),1-19
1147)渡辺勘次(編)(1934) 蚕試彙 (45),21-36
1148)渡辺勘次(1934) 蚕試彙 (46),1-27
1149)渡辺勘次(1935) 蚕試彙 (47),1-38
1150)渡辺勘次(1939) 蚕試彙 (53),1-29
1151)渡辺勘次(1949) 蚕試彙 (63),10-26
1152)渡辺正男,船田敏夫,窪田 厚 外4名(1968) 日蚕雑 37(3),213-218
1153)山田たけを,山崎 寿,小林あつ子(1966) 日蚕雑 35(3),231
1154)山ロ定次郎(1940) 教育農芸 9(11),28-34
1155)山ロ定次郎(1940) 蚕糸 (381),28-31
1156)山ロ定次郎(1943) 蚕糸雑 14(3),144-147
1157)山ロ定次郎(1946) 蚕糸科学 1(1),5-6
1158)山ロ定次郎(1951) 信大繊維報 (1),12-20
1159)山ロ定次郎(1959) 全国蚕種研究会講習会記録 (4),97-129
1160)山ロ定次郎,市川智子(1952) 日蚕雑 21(2・3),123-124
1161)山ロ定次郎,小林 敏(1939) 蚕糸雑 11(3),199-201
1162)山ロ定次郎,小林 敏(1940) 日蚕雑 11(3),204-205
1163)山ロ定次郎,小林 敏(1941) 蚕糸雑 13(1),28-41
1164)山ロ定次郎,清水 猛(1951) 日蚕雑 20(4),299
1165)山ロ定次郎,清水 猛(1953) 日蚕雑 22(3),137
1166)山本茂樹,肥土四郎,大久保英雄(1954) 埼玉蚕試要報 (27),7-10
1167)山本茂樹,長島栄一,浜田喜久二,田中勇吉(1951) 埼玉蚕試要報 (22),38-40
1168)山下秀雄(1961) 日蚕雑 30(3),290-291
1169)山下興亜(1965) 日蚕雑 34(1),1-8
1170)山下興亜,長谷川金作(1964) 日蚕雑 33(2),115-123
1171)山下興亜,長谷川金作(1964) 日蚕雑 33(5),407-416
1172)山下興亜,長谷川金作(1965) 日蚕雑 34(4),235-243
1173)YAMASHITA,O. and HASEGAWA,K.(1966) Jour.Ins.Physiol. 12,325-330
1174)YAMASHITA,O. and HASEGAWA,K.(1966) Jour.Ins.Physiol. 12,957-962
1175)山下幸雄,谷喜代子,小林勝利(1961) 蚕糸研究 (39),12-15
1176)山崎 寿(1954) 日蚕雑 23(6),357-365
1177)山崎 寿(1955) 長野蚕試報 10(3),1-242
1178)山崎 寿(1966) 蚕病に対する先覚者小岩井宗作-その蚕種消毒論および養蚕法について.蚕の作柄をより良くするの会
1179)山崎 寿,小林あつ子,山田たけを(1966) 日蚕雑 35(3),231
1180)山崎 寿,山田たけを,小林あつ子(1965) 日蚕雑 34(5),355-358
1181)山崎 寿,山田たけを,小林あつ子(1966) 日蚕雑 35(3),161-164
1182)柳田頼母(1927) 蚕糸 15(215),22-28
1183)柳田頼母(1928) 蚕卵胚子発育図説.明文堂,東京
1184)梁池三千雄(1964) 熊本蚕試桑と蚕 (7),35-37
1185)安村作郎(1965) 日蚕雑 34(3),205
1186)横川正一(1964) 日蚕雑 33(3),238-239
1187)横川正一(1964) 日蚕雑 33(5),394-398
1188)横川正一(1966) 埼玉蚕試要報 (38),68-72
1189)横川正一,諸口会三(1965) 日蚕雑 34(3),204
1190)横川正一,岡部房之(1960) 日蚕雑 29(2),125-128
1191)横川正一,山口邦友,岡部房之(1963) 日蚕雑 32(3),180
1192)横川正一,山口邦友,岡部房之(1963) 日蚕雑 32(3),180-181
1193)横尾多美男,服部徳一(1953) 応動 17(3・4),161-169
1194)横田長太郎,荒木武雄,富樫 健,野中留蔵(1912) 東京蚕講成績 (47),163-223
1195)横山桂馬(1929) 応動 1(3・4),240-242
1196)横山桂馬(1930) 応動 2(3),221-224
1197)横山忠雄(1929) 応動 1(2),71-80
1198)横山忠雄,高島成之(1952) 蚕糸研究 (2),159-164
1199)YOKOYAMA,T. and KEISTER,M.L.(1953) Ann.Entom.Soc.Amer. 46(2),218-220
1200)横山豊重,丸茂節男(1957) 日蚕雑 26(3),221
1201)横山豊重,三沢秀敏,米山光郎(1964) 山梨蚕試要報 (2),25-27
1202)吉田清隆(1954) 鹿児島蚕試年報 (2),22-23
1203)吉武成美(1954) 日蚕雑 23(2),67-75
1204)吉武成美(1954) 日蚕雑 23(6),343-348
1205)吉武成美(1954) 日蚕報 23(6),349-356
1206)吉武成美(1955) 日蚕雑 24(2),108-113
1207)吉武成美(1956) 日蚕雑 25(6),434-438