
 |
 トレハロース合成はなぜ誘導されるのか? トレハロース合成はなぜ誘導されるのか? |
| LEAタンパク質とクリプトビオシス |
| トレハローストランスポーターの発見 |
| ネムリユスリカの放射線耐性 |
| 参考文献 |
体から水分が抜けていくと、体内の糖類やイオンなどの溶質の濃度が相対的に増加し、浸透圧の上昇が起こります。この浸透圧の変化に着目して、体内浸透圧を擬似的に変化させる実験を行いました。いろいろな濃度の塩化ナトリウム、マンニトール、グリセロール、DMSO(dimethyl sulfoxide)水溶液で幼虫を24時間浸し、その後の体内のトレハロース含量を測定しました。その結果、塩化ナトリウム水溶液では約350ミリオスモル(約1%の濃度)をピークにトレハロースの蓄積がみられました。それは乾燥時のトレハロース蓄積量に匹敵する量でした。これに対してほかの処理区では、浸透圧を上昇させてもトレハロースの蓄積は見られませんでした。

縄文時代の遺跡から発掘されたハスの種が約2000年の時を経て発芽した、という例もあるように、植物の種子は一般的に乾燥に強く、休眠状態にあるともいえます。この種子が休眠に入っていく過程で、大量に合成蓄積されるタンパク質が約30年前に報告され、LEA(late embryogenesis abundant)タンパク質と名付けられました。同じく乾燥耐性のある花粉の中にも大量に蓄積されることから、乾燥耐性に関連したタンパク質と考えられています。その後、葉や根など植物全体においても、乾燥や塩ストレス(イオン濃度の変化)によってLEAタンパク質は蓄積されることがわかりました。
LEAタンパク質は、その一次構造と発現パターンの違いからGroup1~6に分類されていますが、共通して高い親水性(水と結合しやすい)をもっています。試験管内での実験では、乾燥や塩ストレスなどに伴うタンパク質の凝集を抑える効果があることが確認されています。また、一般的なタンパク質が乾燥ストレスによって構造が変化するのに対して、LEAタンパク質では、通常はランダムな状態で構造を持たず、乾燥ストレスを加えるとα-ヘリックスのコイル状に構造化する、という興味深い性質を持っています。

植物に特異的であると思われていたLEAタンパク質ですが、2002年にTunnacliffe博士らによって、クリプトビオシスをする線虫にも存在することが報告されました。つまり、動物にも存在するタンパク質であることが発見されたのです。その後我々のグループも、ネムリユスリカ幼虫からGroup3に属するLEAタンパク質をコードする3つのcDNA(PvLea1、PvLea2、PvLea3)を単離することに成功しました。それからバキュロウィルスを用いてタンパク質を合成し、15分間煮沸したところ、沈殿凝集は見られませんでした。そのことからネムリユスリカの3つのLEAタンパク質も高い親水性を持つことが明らかになりました。また、3つの遺伝子とも乾燥および塩ストレスにより誘導されることがわかりました。
これらのことから、トレハロースの場合と同様、乾燥に伴う体液中のイオン濃度の変化が引き金になって、ネムリユスリカのLEAタンパク質の遺伝子発現が誘導されるものと考えられます。
ではLEAタンパク質はネムリユスリカが乾燥状態になる際、どのような働きをしているのか、仮説を立ててみました。大きく分けて二つの働きが考えられます。
一つ目として、幼虫の乾燥が進むにつれ、ランダム状だったGroup3LEAタンパク質はαヘリックス構造へと変化していきます。このヘリックス構造の親水表面はプラスとマイナスに分極しているめ、細胞内のイオンを結合することによって他のタンパク質が塩析(高濃度の塩によりタンパク質が凝集など変性すること)することを防いでいる可能性があります。またLEAタンパク質は両親媒性であるため、生体膜やタンパク質の疎水性の部分と結合して、生体膜やタンパク質の不可逆的な変性を防いでいる可能性があります。
二つ目として、Group3LEAタンパク質は、ヘリックスの疎水表面同士が結合して立体構造を作ります。これが乾燥に伴い繊維化が進んだ結果、細胞骨格のようになり、あたかも鉄筋コンクリートの鉄筋部分のようにガラス化したトレハロースの構造を強化して、乾燥に伴う細胞の過剰な収縮を防いでいる可能性が考えられます。

これらのことから、LEAタンパク質は、乾燥に伴い爆発的に増加し、トレハロースと協調しながらクリプトビオシス状態の生体成分を保護していると考えられます。
細胞はリン脂質の二重膜で囲まれた袋状の構造をしています。このリン脂質二重膜は水に溶けた物質(たとえば糖、アミノ酸やイオン)をまったく透さないどころか、水自体もほとんど透過することはありません。
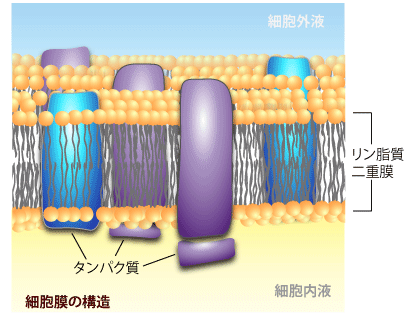
では普段、どのようにして水や水溶性の物質が細胞内外を行き来しているかというと、リン脂質二重膜に埋め込まれたトランスポーターやチャネルと呼ばれる、膜タンパク質を経由して移動しています。このトランスポーターは様々な種類があり、透過する物質の選択性の高いものから、幅広いものまで存在します。たとえば人間の場合、どんなに食事を摂ったとしても、グルコーストランスポーターがなければ、血液中のグルコース(ブドウ糖)が体中の各細胞の中にいきわたることは出来ません。
ネムリユスリカの場合も、脂肪体で大量に合成されたトレハロースを個々の細胞に行きわたらせるには、トレハローストランスポーターが存在することが予想されました。しかし、これまで多細胞生物からトレハローストランスポーターは同定されていませんでした。そこで、ネムリユスリカにはトレハローストランスポーターが多く存在していると予想して、遺伝子の単離を試みました。その結果、完全長のcDNAの同定に成功しました。それから合成したcRNAを、本来はトレハロースを持たないアフリカツメガエルの卵母細胞に人為的に入れ発現させたところ、卵母細胞内にトレハロースの存在が確認され、膜がトレハロースを選択的に取り込んでいることがわかりました。このことから、得られたcDNAはトレハロースのトランスポーターをコードしていることが明らかになり、Tret1遺伝子と命名しました。
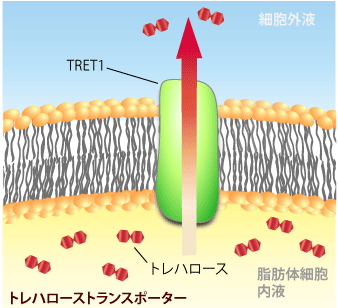
このTret1遺伝子より作成したTRET1タンパク質の性質を調べた結果、トレハロースに対する基質特異性が非常に高く、またその輸送に細胞のエネルギーをまったく必要としない促進拡散型のトランスポーターであることがわかりました。さらにTRET1のトレハロース輸送能力は非常に高く、100mM以上の濃度のトレハロースも十分な速度で透過させることが出来ました。これはクリプトビオシス誘導中のネムリユスリカ体内のトレハロース濃度に匹敵します。またTret1遺伝子は、脂肪体のみで発現が見られ、また乾燥していく過程で発現量が増大していることがわかりました。
これらのことから、TRET1はネムリユスリカ幼虫の脂肪体内で爆発的に合成されるトレハロースを、効率よく体液側に放出する役割を持っていることが示唆されました。
ネムリユスリカは、乾燥していない幼虫(以後活動幼虫とします)よりも、乾燥幼虫のほうが高い放射線量を浴びても生存することが出来ます。ここでの「生存」とは、放射線照射から48時間後に生きている、ということです。しかし、その後の発育や生殖について調べていくと、面白い事実が見えてきました。100Gyのγ線を照射した後、生存したものを飼育していくと、活動幼虫では次世代を残せるが、乾燥幼虫では残せない、ということが判りました。メス成虫は産卵はするものの、その卵は孵化しないのです。生殖腺に影響が出ない線量を比べても、活動幼虫30Gy、乾燥幼虫10Gyと、乾燥幼虫のほうがより放射線の影響が大きいことがわかります。生き物にとって次世代を残す、ということは生存する大きな目的のひとつです。それに関わる生殖腺の放射線からの保護という観点では、乾燥することは負の効果を持つことが示唆されました。

一般的に、放射線耐性を持つ生物は乾燥にも強いことがわかっています。しかし、ここでいう「耐性」とは、自然界には存在しない放射線量に対するものです。遭遇する可能性のないものに対して、生物がその耐性を獲得する方向に進化してきたとは考えられません。おそらく、乾燥に耐えるために発達してきた能力が、放射線に対する耐性と重なっていると考えられます。
そもそも、放射線による影響とはどのようなものなのでしょうか。放射線は、その種類によって組織に与えるエネルギーが異なり、これを線質といいます。線質には大きく低LET放射線と、高LET放射線があり、低LET放射線にはレントゲンで使われるX線やγ線、高LET放射線には最新医療などで使われ始めた陽子線やイオンビームなどが含まれます。これら放射線が生体に吸収されそのエネルギーが細胞内の分子や原子を電離させたりするのですが、それが標的そのものに起きた場合を直接作用、水を介して起きた場合(水が放射線により、イオン、フリーラジカルなどの不安定物質を生じ、それが標的を攻撃する)の間接作用があり、低LET放射線は間接作用、高LET放射線は直接的作用が主体となっています。これらの作用によりDNAが最も影響を受けやすく、一本鎖切断や二本鎖切断を引き起こす原因となっています。


細胞が乾燥に至る過程でもラジカルが発生して、DNAや細胞膜を傷つけています。トレハロースやLEAタンパク質にはそれらを補足する作用があると考えられていて、低LET放射線からのダメージを軽減する一因となっています。しかし、ネムリユスリカについて考えた場合、乾燥幼虫はγ線、そしてDNAを直接切断するイオンビームのいずれにも明らかな耐性を示していることから、DNA修復能が高いのではないかと考えました。実際、DNA修復能のひとつの指標となる、Dq(準いき値線量、放射線のダメージから回復する能力の指標)を比べてみました。普通種であるセスジユスリカとヤモンユスリカ、そしてネムリユスリカの活動幼虫と乾燥幼虫で比較したところ、ネムリユスリカのほうがほかの2種よりもDq値が高く、先天的にDNA修復能が高いことが示唆されました。さらに、乾燥している状態では、Dq値がより高くなることがわかりました。
現在では、乾燥状態を誘導する際に、DNA修復酵素の発現が増加することがわかってきています。さらに、活動幼虫と乾燥幼虫との中間的な生理状態の幼虫にイオンビームを照射したところ、乾燥状態に近いものほど長生きできることがわかりました。この結果から、乾燥状態に移行する過程で、徐々にDNA修復能が上昇していると考えられます。

ここで気をつけていただきたいのは、これは本来、乾燥状態から身を守るために進化した形質であるから、乾燥状態を保ち続けその後蘇生するために必要である、ということです。つまり、乾燥した細胞が無傷でないがゆえに必要な形質である、ということを示唆しています。実際、蘇生させた直後の細胞のDNAの損傷状態をコメットアッセイという方法で調べたところ、ほとんどの細胞のDNAは切断されていました。今まで、ネムリユスリカが乾燥状態でも死なずに生存し続けるということは、細胞は無傷のまま保たれる、と考えられてきました。しかし、やはり脱水は相当なストレスであり、細胞のDNAは損傷を受けていました。ただ、ネムリユスリカにとってこの損傷は、あらかじめ準備されたDNA修復酵素によって修復できる範囲の傷である、ということが考えられます。